
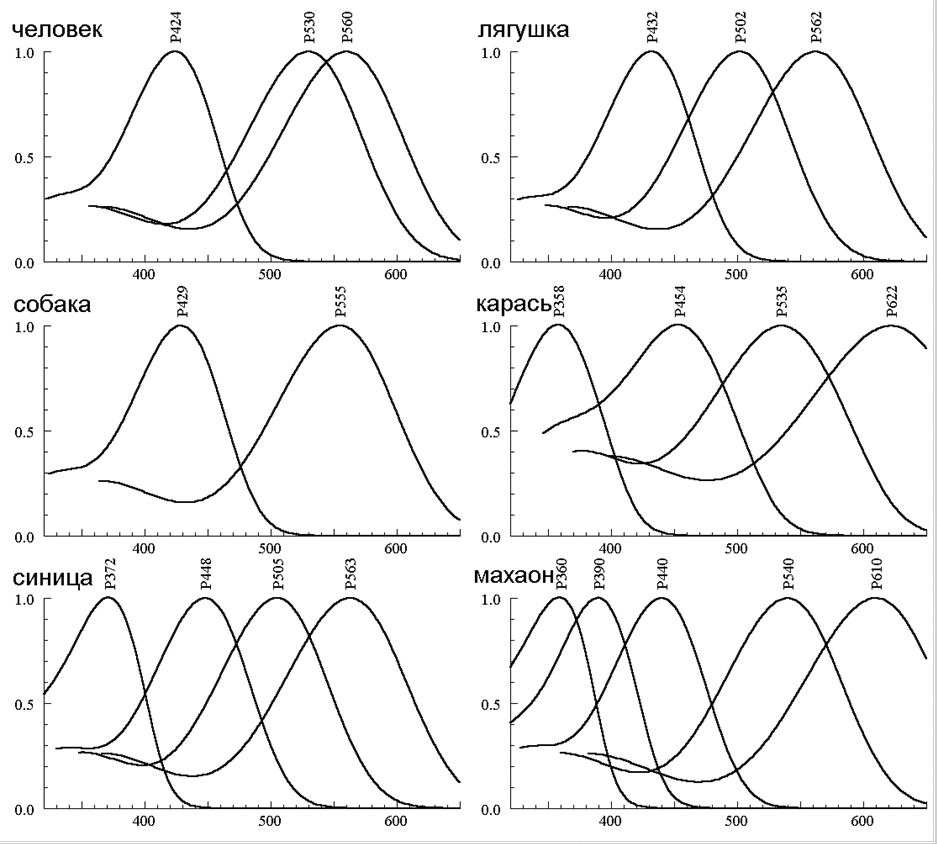
Каждый человек представляет себе, что такое цветовое зрение днем и замечает его утрату в сумерки, когда видимый мир теряет краски. Днем, при высоком уровне освещения, зрение человека обеспечивается работой трех приемников: колбочек, чувствительных к длинноволновой (L), средневолновой (M) и коротковолновой (S) областям спектра. При низкой освещенности (в сумерки, ночью) все три типа колбочек перестают работать, и им на смену приходят палочки — рецепторы с бо́льшей абсолютной чувствительностью (которые днем не работают) (рис. 1а). Все палочки обладают единой спектральной чувствительностью, т. е. представляют собой один приемник. Человек с одним работающим приемником становится ахроматом.
Строгое определение цветового зрения (ЦЗ) — это способность различать излучение по длине волны, независимо от интенсивности. Для этого необходимо (1) иметь в сетчатке не менее двух приемников — двух типов рецепторов с разной спектральной чувствительностью и (2) иметь возможность сравнивать сигналы этих рецепторов. Существует гипотеза, что механизм сравнения сигналов соседних рецепторов (латеральное торможение), полезный для подчеркивания границ, выделения контуров предметов, существовал в глазах, формирующих изображение, до появления в рецепторах двух разных зрительных пигментов [1]. Есть и другая точка зрения, что механизм сравнения длин волн не требует хорошей оптики и мог бы существовать и до появления глаз, формирующих изображение [2]. Авторы этих двух гипотез едины в одном: ЦЗ у позвоночных возникло более 500 млн лет назад и с тех пор поддерживается эволюцией. У беспозвоночных ЦЗ возникло еще раньше [3].
Животное с одним типом рецепторов в сетчатке (с одним приемником) способно различать излучение лишь по яркости. Такое животное, повторим, называется ахромат. Животное с двумя типами рецепторов (с двумя приемниками) называется дихромат. В животном мире известны и три-, и тетра-, и пентахроматы. «Чемпионами» по количеству приемников являются раки-богомолы: у видов, живущих на коралловых рифах в условиях хорошего освещения до 12 приемников (рис. 2) [4]!


Рис. 2. Рак-богомол и кривые спектральной чувствительности его приемников
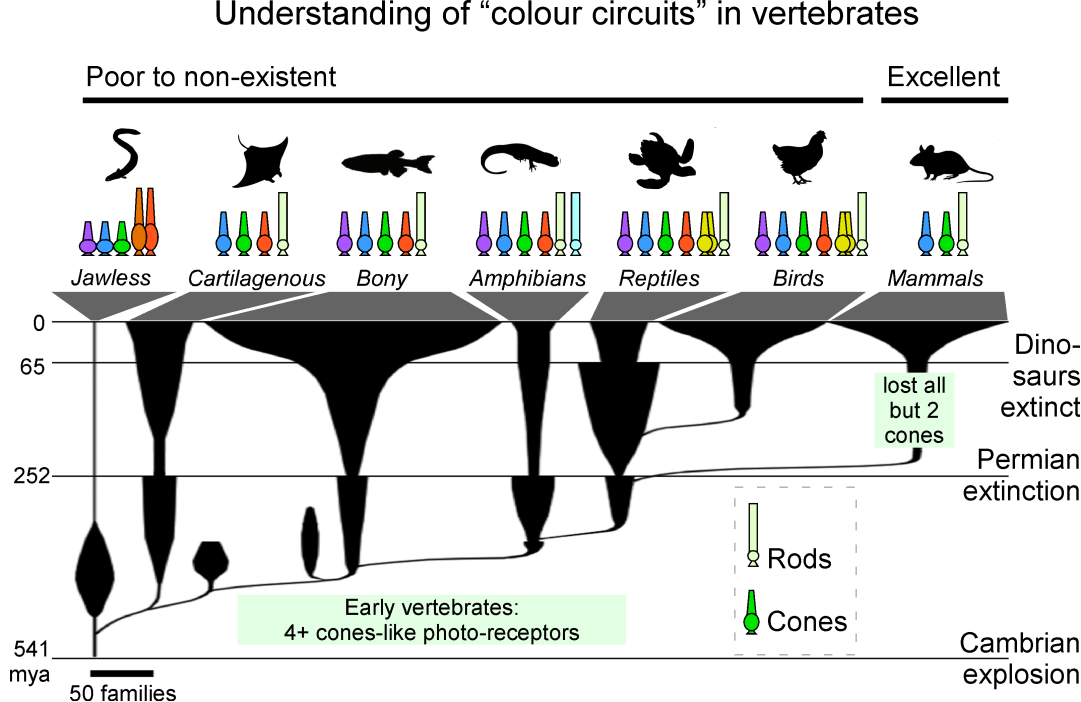
Современное представление о ЦЗ в мире позвоночных животных, времени его возникновения и эволюции прекрасно иллюстрирует эволюционное древо Бадэна (рис. 3) [5].
О рецепторах и зрительных пигментах
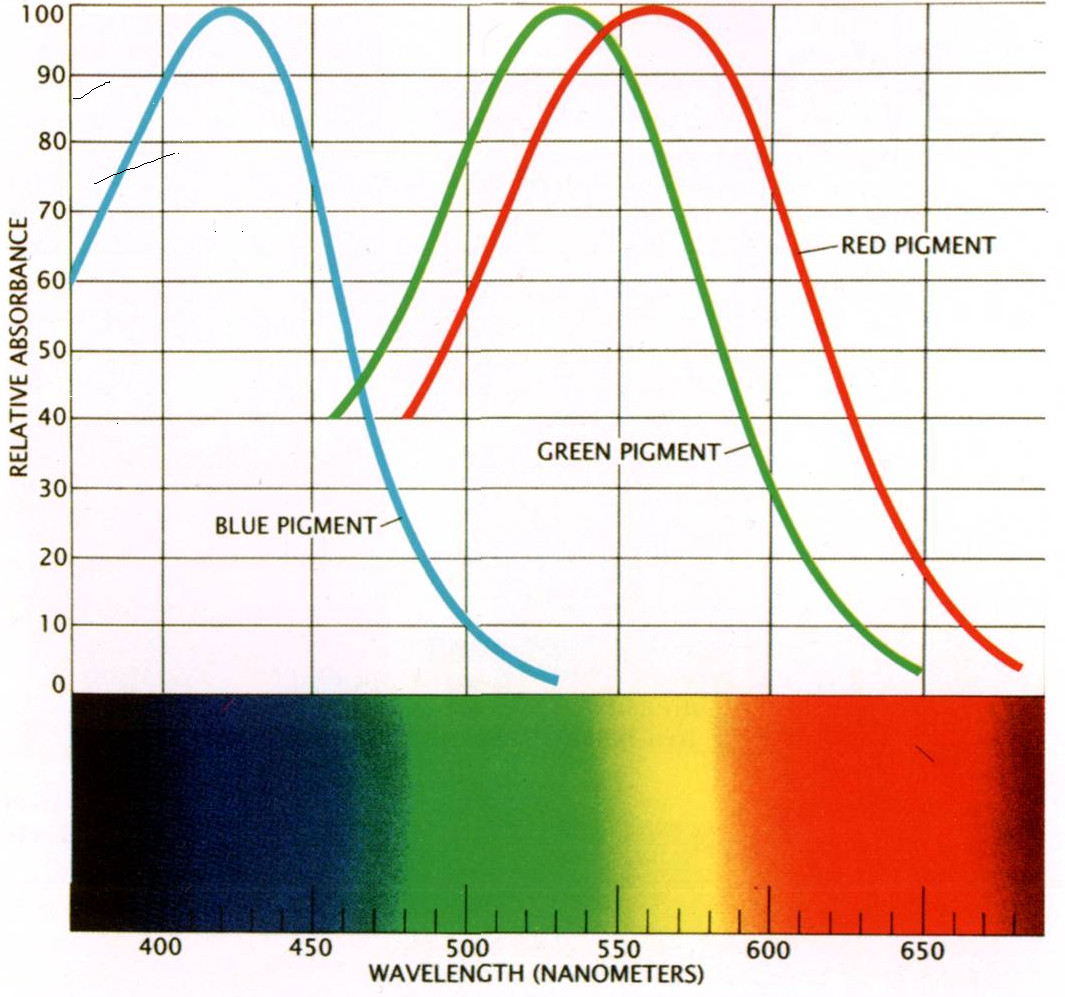
Спектральную чувствительность рецептора (колбочки, палочки) определяет зрительный пигмент, встроенный в мембрану наружного сегмента рецептора. Зрительный пигмент — это комплекс белка (опсина) и альдегида витамина А (хромофора). Опсины фотопигментов кодируются специальными генами. Существуют четыре группы опсинов. Они произошли в результате дупликации гена предкового опсина позвоночных и последующих мутаций более 500 млн лет назад. У современных рыб, рептилий и птиц опсины принадлежат к четырем предковым группам опсинов позвоночных. Молекулярная структура опсина + хромофор определяет форму и положение максимума кривой спектральной чувствительности зрительного пигмента. Положение максимума чувствительности зависит от аминокислотных замен в нескольких «чувствительных» сайтах молекулы опсина и типа хромофора. Хромофор зрительных пигментов морских и наземных позвоночных — это альдегид витамина А1, а хромофор пресноводных рыб — альдегид витамина А2. Колоколообразные кривые пигментов, содержащих в качестве хромофора альдегид витамина А2, сдвинуты в красную область спектра на ~50 нм относительно содержащих альдегид А1 (рис. 1).
Особенности рецепторов ящериц, черепах и птиц
Методом микроспектрофотометрии установлено, что у большинства птиц имеется четыре спектрально различных типа одиночных колбочек, содержащих пигменты с максимумами чувствительности 570, 505, 440 нм (или около 420 или 365 нм в ближнем УФ). Однако спектральная чувствительность колбочек птиц, черепах и ящериц определяется не только фотопигментами, но и встроенными в рецепторы светофильтрами [6]. Три длинноволновых спектральных класса колбочек содержат прозрачные, окрашенные каротиноидами, красные, оранжевые и желтые жировые капли (рис. 4). Эти органеллы находятся на пути света к наружным сегментам колбочек и действуют как режущие (cut-off) фильтры — поглощают («режут») свет короче 572 и 514 нм и таким образом сужают кривые спектральной чувствительности колбочек и смещают их максимумы в длинноволновую область спектра [6, 7]. В коротковолновых колбочках жировые капли бесцветны.
 Схема расположения окрашенных каротиноидами жировых капель. б) Вид изолированной сетчатки черепахи под микроскопом со стороны рецепторов")
 Схема расположения окрашенных каротиноидами жировых капель. б) Вид изолированной сетчатки черепахи под микроскопом со стороны рецепторов")
а) Схема расположения окрашенных каротиноидами жировых капель.
б) Вид изолированной сетчатки черепахи под микроскопом со стороны рецепторов
Можно наблюдать в микроскоп на изолированной инвертированной сетчатке черепахи или ящерицы, как сначала прозрачные красные, оранжевые и желтые (в белом свете) шарики при изменении спектра освещения препарата сетчатки (на просвет) монохроматическим светом от красного к синему становятся темными (непрозрачными): сначала темнеют красные, потом оранжевые, затем и желтые. Прозрачными остаются только капли в колбочках, чувствительных к УФ.
У ночных птиц (сов) и пингвинов, охотящихся на глубине свыше 300 м, капли депигментированы. Отсутствуют жировые капли у крокодилов и змей [6].
Впервые внутриколбочковые окрашенные капли были описаны немецкими морфологами Шульце и Краузэ почти двести лет назад. Эти авторы и предложили гипотезу о возможной роли этих органелл в организации ЦЗ.
Проявление цветоразличительных способностей в поведении разных животных в естественных и экспериментальных условиях
Набор разных фоторецепторов в сетчатке и наличие в зрительной системе нервных клеток, сравнивающих их сигналы, так называемых цветооппонентных клеток, — это необходимая «элементная база» для осуществления у животного ЦЗ. Интересно наблюдать, как и при каких обстоятельствах ЦЗ используется животным, в каких формах поведения. Последнее уточнение может показаться надуманным. Однако на жабах и лягушках показано, что ЦЗ они используют в брачном поведении и не используют в пищевом. В брачный период самцы прудовой лягушки выбирают красные модели и даже предпочитают их натуральным самкам: красная пластмассовая мыльница, которую на веревочке тянет экспериментатор через апрельский прудик, через минуту облепляется самцами, забывшими о самках. Самцы жаб при выборе из красных и голубых моделей предпочитают голубые. В пищевом поведении и лягушки, и жабы цветно слепы [8].
Проявление ЦЗ у рептилий особенно заметно тоже в брачный период, когда самцы агам, вскочив на возвышение, раздувают ярко окрашенный (желтый, красный) подглоточный мешок, привлекая самок и заявляя другим самцам о своем территориальном праве; самцы хамелеонов, встретившись, останавливаются и «полыхают красками», и менее яркий вскоре совсем тускнеет и уходит «с соревнования» без боя.
Пустынная ушастая круглоголовка имеет охристую защитную окраску, но в позе угрозы она приподнимается на передних лапах, скручивает кольцом полосатый хвост и широко разевает ярко-красную пасть.
Не вызывает сомнений, что у дневных птиц, поведение которых во многом зрительно обусловлено, прекрасное ЦЗ. Садоводам известно, как птицы внимательно следят за созреванием плодов желтой облепихи, оранжевой рябины, красной черешни и вишен. Чуть зазевается хозяин — и птицы собрали урожай!
Яркое многоцветное оперение и сложные ритуалы в брачный период тоже говорят о вовлечении ЦЗ. Для нас скрыты многие рисунки птичьего оперения, различаемого самими птицами, благодаря наличию у них (и отсутствию у нас) УФ-приемника (рис. 1). Птицы используют УФ-чувствительность при выборе брачного партнера. Мы считаем, что у многих видов птиц нет полового диморфизма [9], скажем, не можем отличить самку скворца от самца. А самка скворца видит на шее самца «ожерелье», отражающее УФ. Чем оно ярче — тем привлекательнее самец. Блеск «ожерелья» свидетельствует о безупречной структуре перьев, а значит, и об отсутствии клещей-паразитов.
Самцы шалашников Австралии в брачный период строят из травы и веточек, скрепляя их слюной, беседки (шалаши), вымазывая их изнутри мякотью фруктов, глиной. Это не гнезда, а именно павильоны, беседки для ухаживания за самками. Эти строения и подходы к ним (avenue) они украшают ракушками, камушками, цветами, яркими крылышками насекомых, а в последнее время — яркими пластиковыми пробками от бутылок и прочим ярким цветным мусором с помоек соседних кафе. И архитектура шалашей, и способы украшений видоспецифичны. Самец атласного шалашника (Ptilonorhynchus violaceus), как правило, выбирает украшения синего цвета. Самец другого вида (Chlamydera nuchalis), с красным хохолком, выбирает красные цветы и пробки.
Самки, окрашенные гораздо скромнее самцов, несколько раз приходят посмотреть на разные шалаши. В первый визит (в отсутствии самца) они рассматривают, сравнивают, оценивают качество строений и украшений, запоминают понравившийся шалаш и уходят строить гнездо. Во второй визит самка направляется прямо к понравившемуся ранее шалашу. Самец танцует, ухаживает за пришедшей самкой, показывает и преподносит ей разные красочные предметы, и она допускает спаривание, а затем уходит на гнездо откладывать яйца и выводить потомство. Самец за брачный период спаривается с несколькими самками.
Исследователи пытаются понять, как эволюционируют у разных видов шалашников окраска оперения и орнаменты всей сцены (дисплея) и отдельных ее элементов с точки зрения привлекательности для самок (учитывая четырехмерное ЦЗ этих птиц и эволюционную историю видов) [10, 11].
Про опыты на птицах
Профессор Орехово-Зуевского педагогического института, орнитолог Елена Николаевна Дерим-Оглу многие годы проводила летнюю студенческую практику в окрестных лесах. Они изучали поведение птиц-дуплогнездников (синиц, мухоловок пеструшек), выкармливающих птенцов. В этот период птицы-родители так увлечены и заняты своим делом, что не боятся присутствия людей. Елена Николаевна заметила, что птицы, гнездящиеся в скрытых местах (дуплах или развешанных для них заранее синичниках и дуплянках), при поиске своего гнезда руководствуются только особенностями окружающей среды и не обращают внимания на акустические, обонятельные и даже визуальные отвлекающие сигналы.
Для изучения разных способностей птиц (ЦЗ, умения считать и т. д.) Е.Н. разработала условно-рефлекторную методику с пищевым подкреплением. Только обучали родителей, а положительным подкреплением им служил хороший аппетит и разинутые рты вечно голодных птенцов. Обучение проходило постепенно. Сначала птиц-родителей приучали к тому, что леток синичника окружен (помечен) экраном определенного цвета (или полосатым, или с определенным количеством полос). Затем обычный синичник заменяли на «двухкомнатный» синичник. Гнездо с птенцами помещали в одну из секций, где леток в «квартиру» с птенцами был помечен знакомым экраном, а леток в пустую секцию — другим, отличным, экраном. Убедившись, что птицы-родители привыкли к новой обстановке и запомнили, что птенцы находятся в секции, помеченной знакомым экраном, экспериментатор в отсутствие родителей перемещал гнездо в соседнюю «квартиру». Птицы продолжали приносить еду в секцию, обозначенную знакомой маркировкой, и только не обнаружив там птенцов, направлялись к другому входу. Птицы обычно даже не обращали внимания на подросших птенцов, выглядывающих через вход, помеченный «неправильным» стимулом. Чтобы набрать нужную статистику, опыты проводили на многих гнездах в течение всего периода выкармливания вплоть до вылета птенцов. Количество прилетов родителей к каждому летку тщательно протоколировалось.
В таких опытах ранее было показано, что птицы используют ЦЗ в этой форме поведения. Чтобы исследовать поведенческое значение (употребления, использования) включения УФ-приемника в цветовом зрении у птиц, в качестве стимулов были использованы белые экраны, специально окрашенные одни белилами, поглощающими УФ (zinc white UV-absorbing stimuli), другие — мелом или свинцовыми белилами, отражающими УФ (рис. 5).

Все птицы довольно быстро научились отличать цвета, поглощающие УФ-излучение, от нейтральных. Для экспериментаторов такие опыты очень трудны, так как для человека эти белые экраны совершенно одинаковы1 [12].
Цветовое зрение млекопитающих. Про собак и кошек
Млекопитающие в процессе эволюции, в конкуренции со строго дневными динозаврами были вынуждены (и были способны как теплокровные) на долгое время перейти к ночному и скрытному образу жизни. В результате были утрачены два опсина (из четырех предковых). Большинство млекопитающих (копытные, хищные, грызуны и пр.) — дихроматы. У них два колбочковых пигмента, коротковолновый (S) и средневолновый (M).
Наличие цветового зрения у кошек, собак и копытных, как уже говорилось, показали специальные исследования рецепторного состава их сетчаток и поведенческие эксперименты. Существование двух типов колбочек в сетчатке собак обеспечивает им двумерное ЦЗ. Важен ли для собак в их жизни цвет как признак для распознавания объектов? Для ответа на этот вопрос сотрудниками нашей лаборатории были поставлены специальные опыты2 [13].
Условно-рефлекторной методикой с пищевым подкреплением собак обучали различать цветные стимулы (специально окрашенные бумажные экраны, прикрывающие крышки кормушек). Экспериментаторы намеренно использовали зрительные стимулы, которые собаки могли различать как по яркости, так и по цветности. Животных сначала обучали различать пару зрительных стимулов, отличающихся одновременно двумя признаками (например, темно-желтый, DY, подкрепляемый, против светло-синего, LB, неподкрепляемого). В тестах комбинация признаков в стимулах была обратной, так что каждый из тестирующих стимулов имел только одну из двух характеристик, которые изначально были объединены в положительный тренировочный стимул. Таким образом, по результатам тестирования легко узнать, какой из признаков (цвет или яркость) использовался собакой при обучении и какой тем самым является более информативным для животного. Если бы обученные собаки последовательно (раз за разом) выбирали тестовые стимулы того же цвета, что и положительный тренировочный стимул, несмотря на разницу в яркости, можно было бы заключить, что цвет для собак является более информативным признаком (сигналом), чем яркость. Опыты проводились на улице днем. В них участвовало восемь дворняжек разного темперамента из питомника. Все собаки всегда выбирали светло-желтый, LY, против темно-синего, DB [13].
Издавна было известно, что кошек очень трудно научить различать цвета (если вообще возможно). При условно-рефлекторной методике, чтобы добиться дифференцировки стимулов по цвету, требовалось более полутора тысяч сочетаний. На этом основании делался вывод о второстепенном значении цвета в жизни кошки. Однако кошки бывают разные (как и экспериментаторы). В аналогичных опытах 1980-х годов попалось две кошки и кот, которые научились различать по цвету стимулы уже менее, чем за 400 проб (но одну кошку вообще не удавалось этому научить). Но главное, что в этих опытах продемонстрирована способность кошки отличать специально подобранные такие лиловые стимулы от зеленых, которые не отличаются для дихроматической системы (два приемника кошки обеспечить различение этих цветов не могут) [14]. Это указывает на то, что кошки при решении задач на цветоразличение активно использовали третий, зеленочувствительный приемник, возможно, палочки [14]. К сожалению, эти опыты не удалось продолжить.
Приматы — трихроматы
Долгое время считалось, что единственными трихроматами среди млекопитающих (в большинстве дихроматов) являются приматы: узконосые обезьяны Старого Света (в том числе высшие обезьяны и человек) и плосконосые обезьяны Нового Света рода Aluatta. У дневных приматов трихромазия возникла вторично за счет дупликации гена средневолново чувствительного пигмента. Как показывают молекулярные часы, у приматов Старого Света это произошло 40 млн лет назад, а у ревунов (рода Aluatta) — крупных приматов Нового Света — 14 млн лет назад (Hunt et al., 1998) [15]. Эти два независимых события, разнесенные по времени и территориально, привели к возникновению двух одинаковых наборов из трех зрительных пигментов с максимумами 430 нм, 532 нм и 563 нм для S, M и L колбочек, что говорит, в частности, о жестком давлении естественного отбора на ЦЗ [16].
Сумчатые-трихроматы

Лет двадцать тому назад было открыто трихроматическое ЦЗ у сумчатого животного — поссума-медоеда (Tarsipes rostratus) [17]. Это маленькое создание (не больше мышонка) питается нектаром и пыльцой цветов банксии (кустарник с ярко-желтыми и розовыми цветами — эндемик Австралии) и является основным ее опылителем. В его сетчатке кроме двух колбочковых пигментов, характерных для млекопитающих-дихроматов, обнаружен пигмент, чувствительный в УФ-области спектра (рис. 6). Вскоре оказалось, что среди сумчатых есть еще несколько трихроматов: насекомоядная жирнохвостая сумчатая мышь (Sminthopsis crassicaudata), короткохвостый кенгуру (квокка, Setonix brachyurus) и малый бандикут (Isoodon obesulus) [17]. Все другие сумчатые (кенгуру и др.) — дихроматы.
Глядя на наборы рецепторов с разной спектральной чувствительностью у разных животных, мы понимаем, что все животные видят мир по-своему, по-разному, в разных спектральных диапазонах, не так, как мы (рис. 1, 2, 3). Мы не видим в далеком красном диапазоне (длиннее 700 нм), а пресноводные рыбы видят там прекрасно. Мы не видим УФ (у нас нет УФ-чувствительных колбочек). У мышей, многих рыб (у мальков) и птиц, черепах и четырех сумчатых есть УФ-чувствительные колбочки. Морские черепахи часто попадают в рыболовные сети и гибнут. Недавно появилось сообщение, что если сети освещать УФ, то черепахи попадают в них значительно реже: они их видят и обходят [19].
Для нас скрыты сложные окраски многих цветов, рисунков на крыльях бабочек. Однако хитроумные люди научились, используя приборы, визуализировать УФ-лучи. Можно отличить картины старых мастеров от современных подделок, сфотографировав картины в УФ-лучах на пленку, чувствительную к УФ. Известно, что художники прошлого использовали свинцовые белила, отражающие УФ. Теперь пишут цинковыми белилами (свинцовые токсичны, вредны для здоровья), которые не отражают, а поглощают УФ. Также можно выявить невидимый простым глазом рисунок на венчиках многих цветков и половой диморфизм у бабочек-белянок. Впервые это сделал и описал в книге «Зрение насекомых» Г. А. Мазохин-Поршняков [18].

ЦЗ отсутствует только у нескольких видов современных позвоночных животных: это 13 видов китообразных, 10 видов ластоногих, 3 вида енотов, ночные грызуны и 2 вида ночных приматов [20]. У них отсутствуют коротковолново-чувствительные S-колбочки. Отсутствие ЦЗ у этих животных — загадка. Одна из гипотез — это адаптация к образу жизни, в частности, к условиям низкого уровня освещения или обитанию в мутной среде [21].

Заключение
Долгое время в обывательском сознании существовало убеждение, что цветовым зрением обладает только человек — «венец творения», а животные цветов не видят, их мир скучно бесцветен. Убеждение оказалось предрассудком, характерным для антропоцентрического мира. Я привела некоторые примеры, говорящие о том, что ЦЗ очень важно в жизни животных и используется ими в разных формах поведения: при поиске пищи, брачного партнера, контроле территории, отпугивании хищника или привлечении особей своего вида. Более того, в мире современного цивилизованного человека (во многом искусственном, созданном самим человеком) отсутствие действия естественного отбора по ЦЗ приводит к тому, что в человеческой популяции существуют дихроматы и аномальные трихроматы, которые неизвестны среди обезьян Старого Света.
Елена Максимова,
канд. биол. наук, вед. науч. сотр. ИППИ РАН
1. Maximov V. Colour Vision in Early Vertebrates. Iugoslav. Physiol. Pharmacol. Acta., Vol. 34, No.2, 343–349, 1998.
2. Baden T. Circuit-Mechanisms for Colour Vision in Zebrafish. Current Biology 31:12 PR807-R820.
3. Parker A. R. 1998 Colour in Burgess shale animals and the effect of light on evolution in the Cambrian. Proc. R. Soc. Lond. B265, 967–972.
4. Marshall N. J. 1988. A unique colour and polarization vision system in mantis shrimps. Nature 333, 557–560. doi.org/10.1038/333557a0
5. Baden T. New review on colour vision in the water. Jun 21, 2021.
6. Matthew B. Toomey and Joseph C. Corbo. Evolution, Development and Function of Vertebrate Cone Oil Droplets. Front Neural Circuits. 2017; 11: 97.
7. Govardovskii V. On the role of oil drops in colour vision. Vision Res. 1983; 23 (12): 1739–1740.
8. Орлов О. Ю., Максимов В. В. Цветовое зрение и поведение амфибий. 1982.
9. M. D. Eaton Human vision fails to distinguish widespread sexual dichromatism among sexually ‘‘monochromatic’’ birds PNAS2005 vol. 102 no. 31 10945.
10. Borgia G. 1985. Bower quality, number of decorations, and mating success of male satin bowerbirds (Ptilinorhynchus violaceus): An experimental analysis. Anim. Behav. 33: 266–271.
11. Animal visual systems and the evolution of color patterns: Sensory processing illuminates signal evolution Endler J. A., Endler D. A. Westcott J. R., Madden J. R., Madden T. R. 2005. Evolution 59 (8): 1795–1818.
12. Derim-Oglu E., Maximov V. Small passerines can discriminate ultraviolet surface colours. Vision Research, 1994, Vol. 34, No. 11, pp. 1535–1539. «Мелкие воробьиные могут различать окраски, по-разному отражающие УФ».
13. Kasparson A. A., Badridze J., Maximov V. V. 2013 Colour cues proved to be more informative for dogs than brightness. Proc R Soc B280: 20131356. dx.doi.org/10.1098/rspb.2013.1356
14. Кезели А. Р., Максимов В. В., Ломашвили Н. И., Хомерики М. С., Цхведиани Н. Г. Об участии зеленочувствительного приемника сетчатки кошки в цветоразличении // Физиологический журнал СССР им. И. М. Сеченова, 1987.
15. Hunt et al. 1998. Molecular evolution of trichromacy in primates. Vis. Res. 38, 3299–3306. doi: 10.1016/S0042-6989(97)00443-4
16. Henriques et al. 2021. Uniform trichromacy in Alouatta caraya and Alouatta seniculus: behavioural and genetic colour vision evaluation. Front Zool. 2021; 18: 36. doi: 10.1186/s12983-021-00421-0
17. Arrese C. A., Hart N. S., Thomas N., Beazley L. D., Shand J. Trichromacy in Australian marsupials. Curr Biol. 2002; 12: 657–660.
18. Мазохин-Поршняков Г. А. Зрение насекомых. — Наука, 1975.
19. Wang J., Barkan J., Fisler S., Godinez-Reyes C., Swimmer Y. 2013. Developing ultraviolet illumination of gillnets as a method to reduce sea turtle bycatch. Biol. Lett. 9:20130383. dx.doi.org/10.1098/rsbl.2013.0383
20. Jacobs G. H. (1993). The distribution and nature of colour vision among mammals. Biol. Rev.68, 413–471.
21. Peichl, 2005 Diversity of mammalian photoreceptor properties: Adaptations to habitat and lifestyle? Anat Rec A Discov Mol Cell Evol Biol. 2005 Nov;287(1):1001-12. doi: 10.1002/ar.a.20262.
1 cool.iitp.ru/projects/posters/meta/
2 iitp.ru/upload/userpage/117/KasparsonDogs.wmv
См. также trv-science.ru/2024/05/ob-issledovanii-zreniya-zhivotnyh/

 (6 оценок, среднее: 4,17 из 5)
(6 оценок, среднее: 4,17 из 5)
В середине восьмидесятых отдыхали всей семьей в кемпинге на Северском Донце. Лежу в «палатке», двери и окно, естественно, настеж. На столике рядом подсолнух. Зрелый, по цвету совершенно однородный. И птички — влетают в дверь, вылетают из окна, и на пролете выклевывают семечки. Развлекуха! Вечером посмотрел — все оставшиеся зерна — пустые! Как так? Как они на пролете различают? Потом понял — зрение в другой области. То ли В УФ, то ли в ИК. Нагреваются пустые и полные, естественно, по-разному. А как излучают — до сих пор не знаю. Буду рад, если подскажите.
«Блеск «ожерелья» свидетельствует о безупречной структуре перьев, а значит, и об отсутствии клещей-паразитов.»
Может быть, блеск ей просто нравится? О чём свидетельствуют красные и синие пробки?