
Изучение зрения человека и животных – традиционная тема лаборатории обработки сенсорной информации Института проблем передачи информации РАН им. А. А. Харкевича. Канд. биол. наук Елена Михайловна Максимова, старейший сотрудник лаборатории (работает в ИППИ свыше 60 лет), рассказывает об истории изучения свойств ганглиозных клеток (выходных нейронов сетчатки) электрофизиологическим методом, а также кратко описывает строение глаза камерного типа, сетчатки, электрические реакции клеток сетчатки. Опыты проводятся на живых взрослых рыбах — удобном модельном животном. Реакции ганглиозных клеток регистрируются экстраклеточно от окончаний их аксонов в главном оптическом центре рыб – тектуме. Обсуждается возможная роль ганглиозных клеток – детекторов признаков в организации разных форм зрительно обусловленного поведения.
Главной причиной «большого взрыва» видообразования в начале кембрия стало появление у древних животных глаз, формирующих изображение. Такую гипотезу предложил в своей книжке «В мгновение ока» британский зоолог Эндрю Паркер на основании изучения коллекции древней фауны известного палеонтологического месторождения — Сланцев Бёрджеса в Канаде [1]. Предметное зрение (а не просто светочувствительность) повлияло на обострение отношений «хищник — жертва», развитие когтей и челюстей у одних и защитных панцирей, покровительственных или отпугивающих окрасок — у других. Предметное зрение привело к развитию и совершенствованию двигательной системы древних животных, развитию полового диморфизма и способствовало появлению животных роющего образа жизни как крайней формы скрытного поведения, избегания хищника и т. д. Русское слово «вид» как нельзя лучше (на уровне языка) поддерживает эту гипотезу. Постепенно и палеонтологи-эволюционисты стали принимать эту гипотезу.
У современных животных доминируют глаза двух типов строения: фасеточные — у беспозвоночных артропод и камерные — у позвоночных животных и головоногих моллюсков. Фасеточный (сложный) глаз состоит из множества единообразных глазков — фасеток, где каждой светочувствительной единице (рабдому) придается своя оптика. Фасеточные глаза иногда покрывают практически всю голову насекомого (например, у стрекозы) и обеспечивают обзор в 360°. Фасеточные глаза впервые описаны у живших в морях раннего ордовика и в кембрийский период хищника аномалокариса (рис. 1) и трилобитов.

Строение глаза камерного типа
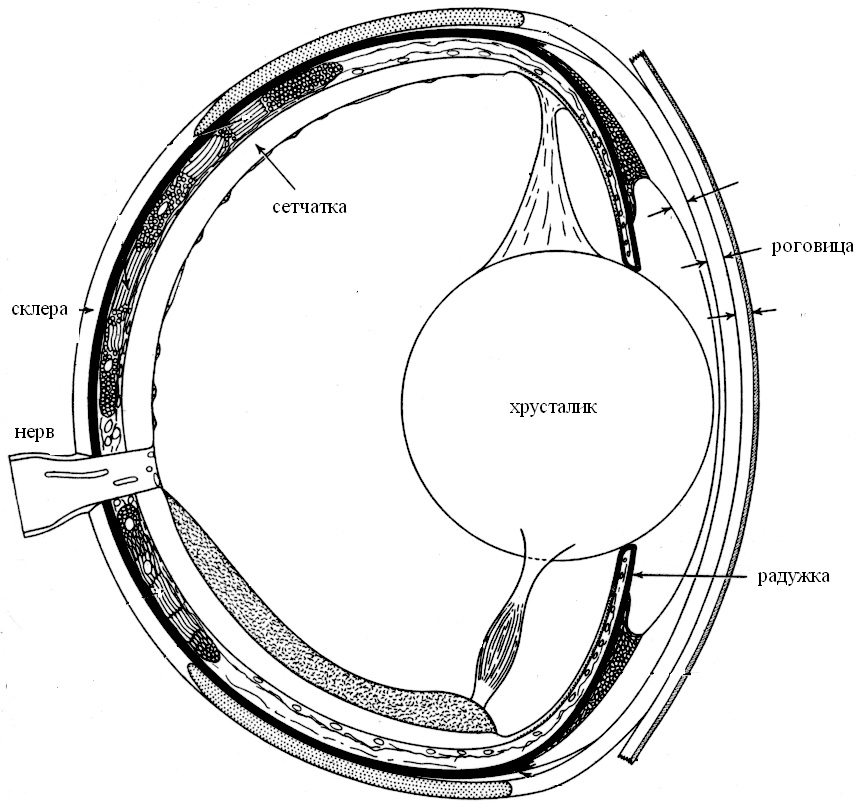
Глаз камерного типа (т. е. подобный фото- или видеокамере) представляет собой устройство (глазное яблоко), где все светочувствительные рецепторы (колбочки и палочки) объединены в единую матрицу — фоторецепторный слой сетчатки. Сетчатка — многослойная нервная ткань, выстилающая заднюю полусферу глаза. Единая оптическая система (роговица, хрусталик) фокусирует на слое рецепторов сетчатки изображение внешнего мира (рис. 2). Два глаза камерного типа располагаются у позвоночных в углублениях черепа (глазницах) фронтально или по бокам головы. Они подвижны в глазницах и благодаря движениям головы обеспечивают достаточно большое поле обзора. На изображении важно различать детали, иначе говоря, иметь высокую остроту зрения, или разрешающую способность. Эта величина обратно пропорциональна угловому расстоянию между рецепторами. Отсюда следует, что хорошо бы иметь большие глаза и очень тонкие рецепторы. У животных с хорошим зрением (птиц, рыб) глаза занимают почти всю черепную коробку.
Развитые глаза камерного типа, вернее, отпечатки такой зрительной системы на черепе описывают у гетеростраков — бесчелюстных, живших на мелководье более 500 млн лет тому назад [2]. За долгий период эволюции камерный глаз позвоночных принципиально не изменился. У приматов в общих чертах глаза такие же, как и у миноги — современного представителя бесчелюстных.
Двумерное инвертированное уменьшенное изображение внешнего мира, проецируемое оптикой глаза на растр наружных сегментов фоторецепторов, каким-то образом трансформируется в представление о его многообразии. Как это происходит? Ответ на этот вопрос пытаются найти морфологи и физиологи, полагая, что знание строения отдельных клеток сетчатки и зрительных отделов мозга, их связей и сигналов, при помощи которых они взаимодействуют, поможет найти ответ на этот вопрос.
Строение сетчатки
Сетчатка камерного глаза позвоночных инвертирована — это значит, что рецепторы расположены в дальнем конце следования светового луча. По дороге к наружным сегментам рецепторов, где находится зрительный пигмент, луч света должен пройти сквозь несколько нервных слоев сетчатки. Клетки сетчатки прозрачны. Существует много способов «проявления» структур сетчатки: разные методы окрашивания, фазово-контрастная микроскопия и др.
В конце XIX — начале XX века великий испанский нейроморфолог Сантъяго Рамон-и-Кахаль исследовал сетчатки разных видов позвоночных животных основных крупных таксонов методом серебрения по Гольджи (произвольное, выборочное окрашивание отдельных клеток со всеми отростками) 1. Он пришел к выводу о едином плане строения сетчаток и конечном основном стандартном наборе ее клеток (фоторецепторы — палочки и колбочки, горизонтальные, биполярные, амакриновые, ганглиозные клетки и глиальные мюллеровские клетки — см. рис. 3) [3].
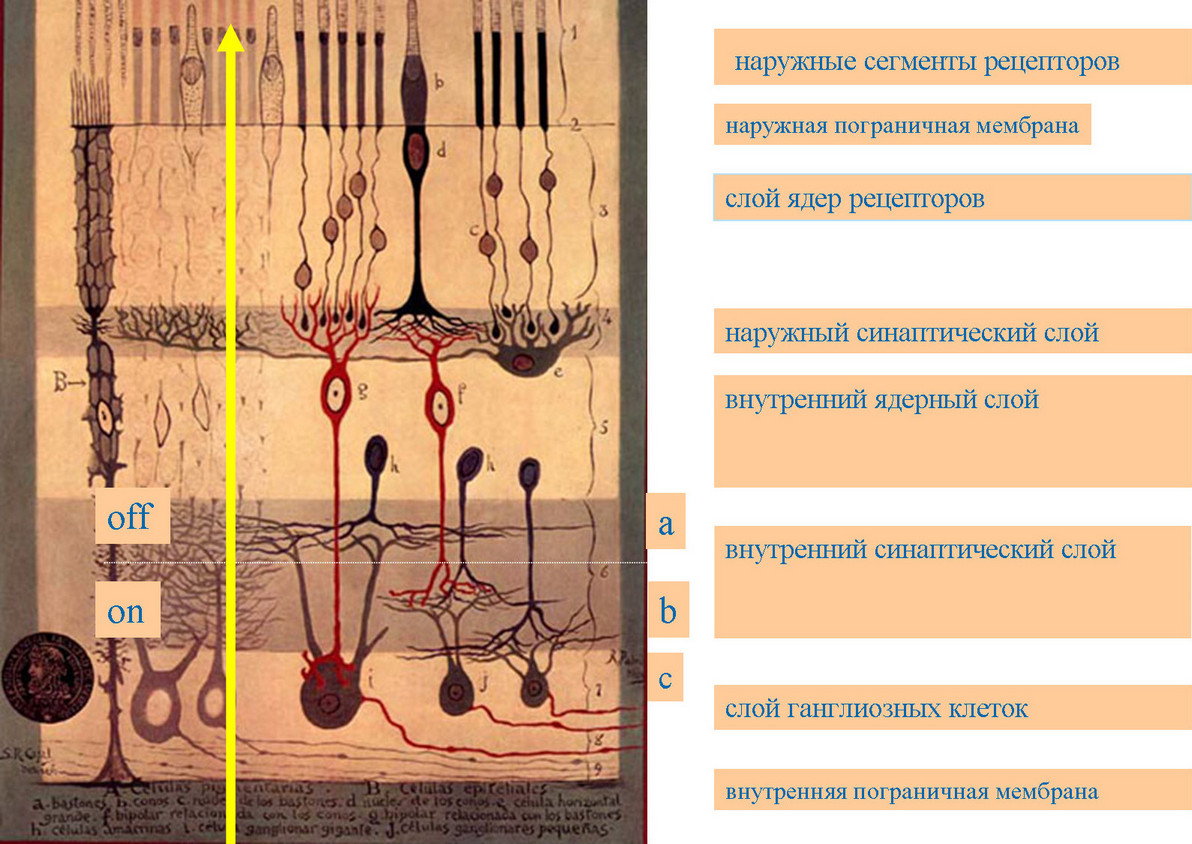
Сетчатка имеет слоистую (экранную) структуру: слой рецепторов, три клеточных (ядерных) и два синаптических (сетчатых) слоя. Синапс — это специализированный контакт между нервными клетками, где происходит передача сигнала от одной клетки к другой, их информационное взаимодействие. В наружном синаптическом слое происходит передача сигналов рецепторов на биполярные и горизонтальные клетки; во внутреннем синаптическом слое аксоны биполяров контактируют с отростками амакриновых клеток нескольких типов и дендритами ганглиозных клеток — выходных нейронов сетчатки.
Р. Кахаль считал, что задача сетчатки — передать точно картинку на растре рецепторов в «умный» мозг. Он находился под впечатлением обнаруженного им такого блока: 1 рецептор — 1 биполяр — 1 ганглиозная клетка. Это — так называемый карликовый путь сигнала в fovea приматов.
Теперь благодаря разнообразным современным методам исследования (электронные микроскопы, фазоконтрастные микроскопы, ультрамикротомы, компьютеризованные конфокальные микроскопы, флюоресцентные красители, иммунохимия, генетические методы, разнообразные трейсеры, регистрация электрических сигналов отдельных клеток сетчатки и популяций клеток) известно не только общее строение сетчатки, но и вид и реакции всех и каждого из ее элементов.
Известны и некоторые узлы морфо-физиологических взаимосвязей этих клеток. Во внутреннем синаптическом слое происходит формирование свойств ганглиозных клеток разных физиологических типов. На основании функциональных, морфологических и транскриптомных данных описано более 40 подтипов ганглиозных клеток сетчатки и 60 подтипов амакриновых клеток сетчатки. По аксонам ганглиозных клеток, составляющих зрительный нерв, их сигналы поступают в зрительные ядра мозга.
Теперь стало понятно, что сетчатка умнее, чем о ней думали раньше, что в ней уже происходит сложная первичная обработка изображения с выделением многих значимых признаков, используемых в дальнейшем нейронами мозга для организации поведения, и справедливо определение сетчатки как части мозга, выдвинутой на периферию [4].
Путь зрительного сигнала
Поглощение кванта света молекулой зрительного пигмента фоторецептора приводит в итоге к изменению электрического потенциала фоторецептора. Возбуждение нервной клетки выражается в ее деполяризации, выделении из ее синаптического окончания нейромедиатора. Он воспринимается постсинаптической мембраной дендрита следующей клетки, что приводит к ее возбуждению. Фоторецепторы беспозвоночных при освещении деполяризуются и генерируют импульсы. Палочки и колбочки сетчатки позвоночных, напротив, деполяризованы в темноте и гиперполяризуются при освещении. Эта удивительная особенность фоторецепторов позвоночных была открыта в 1968 году сотрудником нашей лаборатории Юрием Андреевичем Трифоновым в опытах на изолированных сетчатках рыбы и черепахи и теперь признана всем мировым зрительным сообществом [5].
В синаптических слоях сетчатки сигналы от рецепторов к горизонтальным и биполярным клеткам, от биполярных к ганглиозным клеткам, а также между горизонтальными клетками передаются неимпульсным (аналоговым) способом. Это всё медленные потенциалы, градуально зависящие от силы стимула. Нейромедиатором в цепи передачи сигнала рецептор — биполярная клетка — ганглиозная клетка служит глютамат. Амакриновые клетки — интернейроны внутреннего синаптического слоя — используют множество возбуждающих и тормозных нейромедиаторов, известных в ЦНС. Ганглиозные клетки в ответ на возбуждение формируют ответные (оцифрованные) сигналы в виде разнообразных последовательностей электрических импульсов, которые по аксонам передаются в первичные зрительные центры мозга.
Про модельных животных
Поскольку глаза разных видов позвоночных животных (как говорилось выше) устроены принципиально одинаково, можно выбирать для исследования, в зависимости от задачи, удобное (модельное) животное. Рыбы (как и лягушки) доступны, удобны и в содержании, и в эксперименте как холоднокровные животные. Можно работать как на целом животном, так и на изолированной сетчатке. Последние 30 лет главным модельным животным стала рыбка данио. Она играет роль дрозофилы в генетических работах на позвоночных животных. На прозрачных мальках этой рыбки, интактных или частично обездвиженных, при помощи потенциал-зависимых флуоресцентных красителей (заранее введенных в нервные клетки) регистрируют изменение потенциала возбуждающихся отдельных нервных клеток или их ансамблей (метод Са++-имаджинга) [6]. Картина разноцветной иллюминации отдельных нервных клеток и ансамблей одинаковых функционально клеток в зрительной системе впечатляет и завораживает.
Единство планов строения сетчаток позвоночных позволяет сравнивать результаты исследований, выполненных разными методами разными авторами на разных модельных животных для формирования единой картины о механизмах зрительного процесса и его роли в жизни животных.
Поведение рыб во многом определяется зрением. Врожденные зрительно управляемые формы поведения рыб: фототаксис, оптомоторная реакция, охотничье поведение и оборонительный рефлекс — избегание наблюдаются уже у четырехдневных мальков данио и сохраняются во взрослом состоянии. В организации всех этих разных форм поведения участвует тектум оптикум (ТО) — главный первичный зрительный центр рыб. У рыб в ТО приходит 98% аксонов ганглиозных клеток (ГК).
Мы с 1970-х годов работаем на рыбах электрофизиологическими методами. В настоящее время мы исследуем зрительную систему взрослого карася (близкого родственника данио). Методика регистрации реакций ганглиозных клеток от окончаний их аксонов в ТО в ответ на зрительные стимулы, близкие к реальным, была описана в знаменитой статье «Что глаз лягушки сообщает мозгу лягушки» в 1959 году [8]. Авторы (Матурана, Летвин, Питтс, Маккалох — сотрудники MIT) работали на целой лягушке. Реакции ганглиозных клеток регистрировали не от тел клеток в глазу, а от окончаний их аксонов в тектуме. Лягушка смотрела нормальными неповрежденными глазами на стимулы (темные пятнышки, полоски, границы), которые двигались перед ней по белому экрану. Были специально разработаны низкоомные микроэлектроды (металлические в стекле с электролитически наваренной на конце шляпкой губчатой платины диаметром 2–5 мкм) для экстраклеточного отведения импульсов с амплитудой порядка сотен микровольт от терминальных кисточек аксонов. Импульсы наблюдали на осциллографе и прослушивали через громкоговоритель.
Авторы описали четыре типа реакций, отводимых на разных глубинах в ТО лягушки. Самой выразительной была реакция ГК на подвижное маленькое черное пятнышко. В то же время на включение и выключение света в комнате такие клетки вовсе не реагировали. Такие ГК получили название детекторов малого черного пятна (иногда их называли детекторами жука).
Молодой сотрудник нашей лаборатории Ваня Пигарёв 2, прочитав статью Летвина и его коллег, вскоре первым в нашей стране воспроизвел эту методику (сложнее всего было самостоятельно изготовить специальные микроэлектроды) и получил аналогичные результаты на «советской» лягушке. Потом Пигарёв со своим другом и коллегой Игорем Зенкиным модифицировали методику для работы на рыбах. Работая в командировке в лаборатории физиологии Института внутренних морей в Борке на Волге, они впервые увидели (и услышали) реакции ГК щук, которые реагировали на всё (контрастные границы, пятнышки, полоски), что движется в поле зрения рыбы в направлении от хвоста к голове, и не замечали стимулы, движущиеся в обратном направлении [7]. У лягушки реакции таких клеток в ТО не были описаны. Виртуоз эксперимента, Ваня Пигарёв смонтировал крошечный микроманипулятор (своей конструкции) на черепе свободно плавающей щуки, и все услышали, что мощная реакция этих клеток возникает только на стимулы, движущиеся от хвоста к голове. Коллеги решили, что в природе эти клетки-детекторы движения от хвоста к голове сигнализируют щуке, затаившейся в прибрежной осоке, что ее сносит течением (подводная трава тогда движется от хвоста к голове), и назвали их детекторами сноса. Когда щука движется вперед, стебли растений проплывают мимо глаза спереди назад, и эти детекторы молчат.
Я в то время тоже работала в Борке на изолированной сетчатке щуки с горизонтальными клетками, внутриклеточными электродами. Мощное звучание детекторов щуки из соседней комнаты не давало мне покоя. Я попросила Ваню обучить и меня методике Летвина. С тех пор я так и работаю. С годами совершенствовалась экспериментальная установка. Теперь она оснащена тремя компьютерами, которые генерируют и предъявляют разнообразные стимулы, производят частичную онлайн-обработку реакций, ведут автопротокол и записывают результаты опыта в базу данных. Неизменными остаются только электроды.
В дальнейшем мы с О. Ю. Орловым и В. В. Максимовым изучали зрение разных пресноводных и морских рыб в экспедициях в Институте южных морей в Севастополе и на Звенигородской биостанции. О. Ю. Орлов с учениками работал по этой методике также в Институте моря в ДВНЦ РАН. В течение нескольких лет на базе нашей лаборатории проходил практикум по зрению для студентов биофака кафедры физиологии животных МГУ, где я читала курс физиологии зрения.
Иван Николаевич Пигарёв, будучи в научной командировке в США, навестил Джерри Летвина и порадовал его, рассказав о работах в нашей лаборатории. Группа Джерри Летвина была близка и идейно, и по составу (математики, биологи, морфолог) нашей лаборатории. В 2009 году мы в лаборатории отметили 50-летие выхода статьи Летвина докладом, который сделал на семинаре лаборатории В. В. Максимов.
Выпускник кафедры высшей нервной деятельности Иван Николаевич Пигарёв с работы на щуке вскоре переключился на исследование обработки зрительной информации в корковых и подкорковых структурах мозга ненаркотизированной кошки. Его методика была новаторской, так как весь мир тогда работал на животных под наркозом. Методика безболезненного крепления на черепе (временно, в процессе опыта) отводящей аппаратуры позволяла работать на одном животном месяцами и годами (рис. 4). Он изучал бинокулярное зрение кошки, специфику разных зрительных зон коры, константность восприятия зрительного пространства. В свободное от опытов время кошка жила нормальной жизнью.

В опыте кошка частенько засыпала, закрывала глаза, энцефалограмма демонстрировала картину сна, и на фоне сна иногда возникала активность в зрительных нейронах коры. Этот факт послужил для Пигарёва начальным толчком для разработки висцеральной гипотезы сна 3, которую он увлеченно развивал теоретически и экспериментально до конца жизни [9].
Как проходит опыт на рыбе сейчас
Наша группа осталась верна рыбе. Взрослая рыба — обычно карась — закреплена в специальном станке в аквариуме. Сквозь его прозрачную стенку она смотрит на экран монитора, на котором ей показывают разные генерируемые компьютером картинки: движущиеся пятна, полоски, границы — черно-белые и цветные. Электрод через отверстие в черепе с помощью микроманипулятора под зрительным контролем вводят в ТО, расположенный у рыб поверхностно.
При погружении электрода на разных глубинах мы регистрируем реакции ГК разных типов. Всего таким методом у рыб (не только у карасей, но и у плотвы, карпа, усача, зеленушки, ставриды и др.) в ТО мы с уверенностью определяем реакции тринадцати типов специализированных ГК (возможно, их больше). Это реакции дирекционально избирательных (ДИ) ГК, или детекторов направления движения: первые, на глубине от поверхности 50 мкм реагируют на движение стимулов от хвоста к голове; вторые — на глубине 100 мкм — на движение снизу вверх; третьи — сверху вниз. В каждой группе поровну клеток, предпочитающих движение темных границ по более светлому фону, и наоборот, светлых границ по более темному фону. Итого шесть типов дирекционально избирательных ганглиозных клеток. В том же горизонте глубин можно зарегистрировать детекторы малого белого и черного пятна, похожие по свойствам на детекторы пятна лягушки. Такие клетки не реагируют на выключение и включение общего освещения.
На глубине 150 мкм регистрируются два типа детекторов ориентированных линий (горизонтального и вертикального края). Реакцию первых вызывают подвижные и неподвижные границы и полосы, ориентированные горизонтально, вторых — вертикально. Реакция такой клетки на неподвижный стимул (полоску) предпочтительной ориентации может длиться минутами, но мгновенно прекращается (тормозится) при внесении в поле зрения полоски ортогональной ориентации.
В литературе вышеописанные ГК называют детектрами признаков (контраста, размера, направления движения, ориентации).
Размеры рецептивных полей ГК описанных выше типов составляют от 3° до 5°. Это соответствует области рецепторной поверхности сетчатки, насчитывающей несколько сотен фоторецепторов, с которой ГК получает сигналы. Характер обработки изображения каждой ГК (ее тип), характер ее выходного сигнала определяется специфическими синаптическими связями ее дендрита с нейронами предыдущих этажей (биполярными, горизонтальными, амакриновыми клетками сетчатки). Мозаики дендритов ГК каждого типа (по данным морфологии) полностью покрывают, как кафелем, всю поверхность сетчатки на своем уровне стратификации во ВСС. В ТО окончания ГК, детектирующих разные признаки изображения, образуют ретинотопические карты выделяемого ими признака, каждый тип на своем уровне.
Долгое время наше недоумение вызывал тот факт, что у рыб — животных, обладающих хорошим цветовым зрением, с тремя типами колбочек и разнообразными цветооппонентными горизонтальными и ганглиозными клетками в сетчатке, — мы не обнаруживали соответствующих цветооппонентных реакций в их главном оптическом центре — ТО. Наконец недавно благодаря виртуозному мастерству нашего молодого сотрудника А. Алипера было показано, что ганглиозные клетки с фоновой активностью, аксоны которых образуют самый глубокий и мощный слой в ТО на глубине 200 мкм, и являются этим недостающим звеном. Они делятся на три группы по уровню фоновой активности на свету и в темноте и специфике связей с разными типами колбочек (рис. 5).

Так, соответственно количеству морфофункциональных типов ГК, уже в сетчатке формируются и по аксонам передаются в ТО около двух десятков разных «описаний» картины внешнего мира.
В ТО устанавливается соответствие карты внешнего мира и карты тела.
Набор (комплект) из нескольких карт признаков (таких, как знак контраста стимула относительно фона, его размер, направление движения, ориентация, цвет), расположенных в ТО стопкой друг под другом, составляют ретинотектальную карту значимых признаков (saliency map). Считается, что в ТО по карте значимых признаков происходит выбор главного объекта, видимого в поле зрения (pop-out stimulus), и переключение на него внимания. Известно, что функции фиксации взора на объекте и переключения внимания сохраняются за тектумом и у млекопитающих, у которых уже развивается кора.
Детекторная концепция
Детектор черного пятнышка, описанный в статье «Что глаз лягушки сообщает мозгу лягушки», можно рассматривать как ключевой стимул для запуска охотничьего поведения. В поведенческом опыте лягушка, а лучше жаба, увидев на арене маленькое черное подвижное пятнышко, поворачивается к нему, подходит и выбрасывает липкий язык, пытаясь его съесть. Наблюдение за свободно плавающими (и частично обездвиженными) мальками данио дает представление о том, реакции каких специализированных ГК — детекторов признаков, описанных выше, — используются в поведении. В охотничьем поведении это детекторы малых пятен и ДИ ГК. Объекты, меньшие 5°, интерпретируются как пищевые и вызывают охотничье поведение; объекты, превышающие 10°, вызывают реакцию избегания.
В 1970-е годы тенденция искать в сетчатке детекторы ключевых стимулов поддерживалась и результатами работ Лоренца и Тинбергена по этологии. Первые работы на рыбе обнадеживали. Но постепенно накапливались и опровергающие примеры. У рыб с очень разной экологией, хищных и вегетарианцев, бентосных, пелагических, коралловых, обнаруживались одни и те же наборы детекторов. Детекторы пятна были открыты у кролика и мыши, а детекторы ориентированных линий — не только у рыб и голубей, но и у мышей. Более того, дирекционально избирательные и ориентационно избирательные зрительные реакции обнаружены в соответствующих зрительных ганглиях у дрозофилы и стрекозы 4.
С накоплением данных оказалось, что в сетчатках всех исследованных животных более или менее стандартный набор специализиованных ГК, иногда отличающихся некоторыми деталями. Стало быть, детектирование таких признаков, как знак контраста относительно фона, размер стимула, направление движения, ориентация — это полезные (необходимые) этапы обработки зрительной информации, общие для позвоночных и беспозвоночных животных. (Что не мешает рассматривать, например, детектор пятнышка как ключевой стимул в пищевом поведении рыбы, мухи, цыпленка.)
Итак, нам известно, что сообщает глаз рыбы нейронам главного зрительного центра рыбы. Известно и в каких поведенческих зрительно обусловленных актах принимает участие ТО. Остается понять, узнать, как нейроны ТО организуют эти процессы.
1. Parker A. (2003). In the Blink of an Eye: How Vision Sparked the Big Bang of Evolution. Cambridge, MA: Perseus Pub. ISBN 0–7382–0607–5).
2. Maximov V. V. Environmental factors which may have led to the appearance of colour vision // Phil. T rans. R. Soc. Lond.B (2000) 355, 1239–1242
3. Ramón y Cajal S. (1892). La retine des vertébrés // La Cellule, 9, 119–225.
4. Gollisch T. and Meister M. Eye smarter than scientists believed: Neural computations in circuits of the retina // Neuron. 2010; 65(2): 150–164. doi: 10.1016/j.neuron.2009.12.009.
5. Трифонов Ю. А. Изучение синаптической передачи между фоторецептором и горизонтальной клеткой при помощи электрических раздражений сетчатки // Биофизика. 1968. Т. 13. С. 809–817.
6. Muto A., Ohkura M., Abe G., Nakai J., and Kawakami K. Real-Time Visualization of Neuronal Activity during Perception // Current Biology 23, 307–311, 2013.
7. Зенкин Г. М., Пигарёв И. Н. Детекторные свойства ганглиозных клеток сетчатки щуки // Биофизика. 1969.Т. 14. С. 722–730.
8. Lettvin J. Y. , Maturana H. R. , McCulloch W. S. and Pitts W. H. What the Frog’s Eye Tells the Frog’s Brain. Proc. IRE 47 (1959).
9. Пигарёв И.Н. Висцеральная теория сна // Журнал высшей нервной деятельности. 2013. Т. 1. С. 86—104.
1 Р. Кахаль доказал, что нервная система (в том числе сетчатка) не является непрерывной сетью, как тогда считалось. Она (подобно другим тканям) состоит из отдельных клеточных элементов — нейронов. Нейрон имеет входное устройство — короткий ветвящийся отросток — дендрит, воспринимающий входной сигнал, тело с ядром и выходной отросток — аксон, по которому ответная реакция нейрона распространяется дальше.
2 Иван Николаевич Пигарёв (1941–2021) — автор висцеральной теории сна, докт. биол. наук (1991), сотрудник ИППИ с 1961 года.
3 В бодрствовании нейроны зрительной коры занимаются обработкой сигналов от сетчатки. Во сне они, не получая сигналов от экстерорецепторов, переключаются на обработку сигналов от внутренних органов (желудка, кишок).
4 На первый взгляд отличные друг от друга зрительные системы позвоночных и беспозвоночных имеют общее происхождение, что доказано методами молекулярной генетики (Gehring).
