
Версия видеолекции (youtube.com/watch?v=Z-LRsSpvTeI), адаптированная автором для ТрВ-Наука.
Эволюция старения
Первый сюжет у нас посвящен эволюции старения. На эту тему вышли сразу две интересные статьи. Сначала немного контекста. Эволюция старения — это вообще одна из самых интригующих тем эволюционной биологии. Потому что мы имеем дело с признаком, который, на первый взгляд, вреден. Живым организмам вредно стареть и умирать, это снижает, казалось бы, их дарвиновскую приспособленность, и поэтому отбор должен был бы со старением побороться. Мы знаем, что технически это возможно в каких-то пределах, потому что существуют нестареющие животные. Значит, в принципе эволюция может такое создать. Но тем не менее большинство животных стареет, и это некая научная загадка, привлекающая издавна внимание эволюционистов. Сначала нужно дать определение. Старение — это рост смертности с возрастом. Это простое, удобное и на самом деле грамотное определение. По мере того, как животное становится старше, вероятность, что оно умрет в течение следующего, скажем, года, растет. Причем растет, как правило, экспоненциально. У людей после достижения половой зрелости этот закон четко соблюдается (совсем четко — после 30 лет): вероятность умереть в течение следующего года от всех причин удваивается примерно каждые семь лет.
Я говорил, что есть животные, которые не стареют, но это не значит, что есть бессмертные животные. Бессмертия не бывает, потому что смертность не бывает нулевой. Любое животное может в любой момент погибнуть, потому что его съест хищник, потому что оно заразится смертельным вирусом, оно может умереть от голода и т. д. Смертность не бывает нулевой, но она может не расти с возрастом, и тогда у нас нет старения. Есть даже организмы, у которых смертность снижается с возрастом, и тогда говорят об отрицательном старении.
Старение требует эволюционных объяснений.
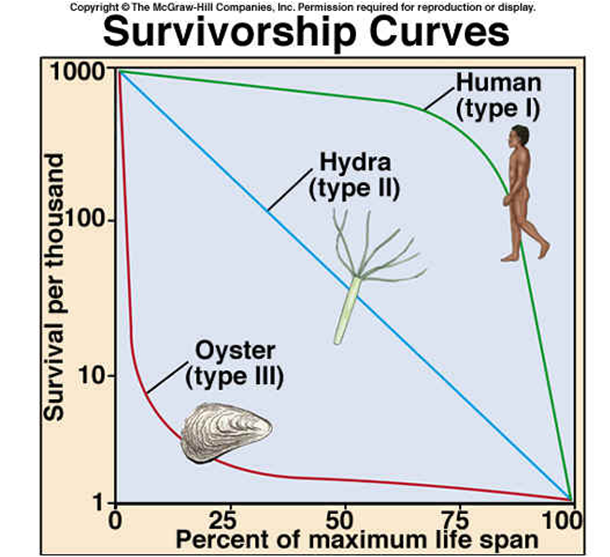
На рис. 1 показаны так называемые кривые выживания — это традиционный способ изображать динамику старения. По вертикальной оси откладываем численность когорты. Когорта — это, например, тысяча животных, родившихся в одно время. Дальше за ними следят, как они постепенно умирают. Здесь по горизонтали отложен возраст, а по вертикали — число доживших. В этих координатах, если старения нет, то график — прямая линия (каждый год умирает, например, один процент). Если наклон растет, как в случае человека, это значит, что старение есть.
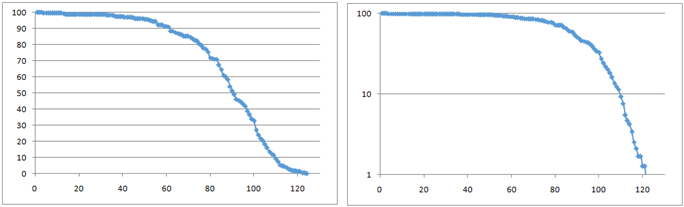
На рис. 2 — реальная линия выживания наших лабораторных дрозофил. Это наши эмпирические данные, когда я работал на кафедре биологической эволюции МГУ. Дрозофилы после выхода из куколки сначала практически не умирают. Они молодые, крепкие, здоровые. Потом, начиная где-то с 40–50 дней, они начинают потихоньку умирать, потом всё быстрее, быстрее. Это значит, что есть хорошо выраженное старение, и должен вам сказать, что линии выживания людей выглядят практически так же, как дрозофильные: такая же характерная выпуклая кривая с сильно выраженным старением.
Почему отбор не убирает старение как очевидно вредный признак?
На эту тему существует огромное количество литературы. Есть две основные группы идей. Первая группа предполагает, что старение — это адаптация, естественный отбор как бы нарочно поддерживает старение, потому что оно чем-то полезно, например увеличивает приспособляемость организмов, ускоряет смену поколений и тем самым помогает популяциям быстрее адаптироваться к меняющимся условиям. Это старая идея, ее высказывал еще Август Вейсман. А в наши дни эту точку зрения очень активно поддерживал академик Владимир Петрович Скулачёв, выдающийся ученый. К сожалению, недавно он скончался в возрасте почти 88 лет. Он считал, что это адаптация — есть генетическая программа старения, поддержанная отбором, и надо ее найти и сломать — тогда мы перестанем стареть. Но на самом деле, по-видимому, сейчас уже можно сказать определенно, что эта идея (старение — это адаптация, «нарочно» созданная естественным отбором) работает только для очень специфических ситуаций и для очень узкого круга живых организмов. Например, это определенно работает для знаменитого модельного червячка Caenorhabditis elegans. Вот у него, судя по всему, действительно старение — это адаптация. Но для большинства животных это объяснение не проходит, к сожалению. Если бы это было так, то со старением было бы гораздо легче бороться. Большинство специалистов считает, что в подавляющем большинстве случаев старение — это не адаптация, а побочный эффект ослабления очищающего отбора с возрастом (я объясню, что это значит) или отбора таких аллелей (генетических вариантов), которые одновременно повышают раннюю приспособленность (например, плодовитость в молодости) и снижают приспособленность в старшем возрасте из-за износа каких-то систем организма. В общем, старение — побочный эффект других эволюционных факторов, и вот эта вторая группа идей называется классической эволюционной теорией старения.
В ее основе лежат две идеи, принадлежащие двум великим биологам. Первая идея принадлежит Питеру Медавару, нобелевскому лауреату. Он рассуждал так. Представим себе, что изначально у нас есть нестареющий вид животных. Но смертность не бывает нулевой. Значит, если старения нет, то всё же особи умирают, просто процент умирающих каждый год будет одинаков, и численность когорты убывает экспоненциально. Будем считать, что они настолько не стареют, что у них даже плодовитость не снижается с возрастом: они отлично размножаются вплоть до самой смерти, сколько бы ни прожили.
При этом ожидаемое количество потомков, которых животное еще оставит в будущем, будет снижаться с возрастом, поскольку чем старше возраст, тем меньше особей до него доживает. Это значит, что вредные мутации, эффекты которых проявляются начиная с какого-то возраста, будут с точки зрения естественного отбора становиться всё менее вредными по мере увеличения этого возраста. Они будут всё слабее уменьшать ожидаемый репродуктивный успех нашего организма, и в конце концов мы достигаем таких возрастов, до которых доживает уже так мало особей, что естественный отбор вообще не сможет заметить, что такие мутации вредны. Таким образом, с возрастом организмы попадают в то, что называют selection shadow (тень отбора). Это такая область, куда уже как бы не проникает очищающий свет отбора. В природе до таких возрастов мало кто доживает. Но если мы возьмем из природы это животное, посадим в лабораторию, где оно будет защищено от хищников, болезней, голода и всех перипетий судьбы, то оно доживет до этого возраста, и тут мы увидим, что оно начинает болеть, слабеть, у него всё начинает разваливаться, и в конце концов оно умирает. Мы увидим старение.
Но одной этой идеи недостаточно. Вторая фундаментальная идея, лежащая в основе классической теории старения, принадлежит другому великому биологу, Джорджу Уильямсу. Больше всего он известен своей книгой «Адаптация и естественный отбор» (1966), в которой Уильямс в пух и прах раскритиковал теорию группового отбора и заложил основы так называемого геноцентрического взгляда на эволюцию — того самого взгляда, который был потом развит и популяризирован Докинзом в книге «Эгоистичный ген». Но сейчас мы говорим о другой идее Уильямса, связанной с эволюцией старения. Эта идея называется страшным словом «антагонистическая плейотропия».
Уильямс обратил внимание, что на самом деле у многих видов особи таки доживают в природе до такого возраста, когда старение уже проявляется. Уильямс предположил, и это предположение потом многократно подтверждалось, что существует множество генетических вариантов, которые повышают приспособленность в молодости ценой ее более быстрого снижения в поздних возрастах. Такие аллели поддерживаются отбором, поскольку ранние признаки важнее поздних — до поздних возрастов доживают немногие. Поскольку ресурсы организма всегда ограничены, если организм очень много ресурсов бросает, например, на раннее размножение, то у него должно оставаться меньше ресурсов на сохранение своего здоровья, на починку накапливающихся повреждений, т. е. он будет быстрее стареть.
Из этих идей Медавара и Уильямса вытекает проверяемое следствие, которое состоит в том, что скорость старения будет зависеть от уровня внешней, не связанной с возрастом смертности. Например, имеется очень сильный пресс хищников — если есть какая-нибудь мышка, которую ловят и едят все, кому не лень, то у такой мышки мало шансов дожить, скажем, до четырехлетнего возраста, даже если она не будет стареть. Всех мышей съедают раньше. Соответственно, мутации, которые портят здоровье мышки в четыре года и позже, будут незаметны для отбора и могут свободно накапливаться. Поэтому такие животные старятся быстро. И совсем другое дело, когда у нас иной зверек такого же размера — летучая мышка, которая умеет летать. Ее не так просто съесть, ведь полет — это великолепная защита от хищников. Соответственно, у летающих животных больше шансов прожить долгую жизнь, и отбор лучше видит поздние вредные мутации. Соответственно, летучие мыши должны стареть медленнее. Это так и есть — действительно, среди рукокрылых много рекордсменов по продолжительности жизни, а обычные мышки живут очень недолго и быстро стареют.
Похожая ситуация с птицами — птицы в среднем стареют гораздо медленнее, чем нелетающие звери такого же размера. Это хорошо согласуется с данным предсказанием. Оно было сделано самим Уильямсом в замечательной статье 1957 года, это одна из самых цитируемых статей в литературе по эволюции старения — много тысяч ссылок.
И вот в прошлом году появились две работы, подтверждающих это предсказание. Одна из них — по амфибиям и рептилиям1. У холоднокровных вообще вопросы старения и продолжительности жизни очень мало изучены, гораздо меньше, чем у птиц и млекопитающих. В этой работе большой авторский коллектив собрал огромный массив данных по 77 видам амфибий и рептилий из 17 природных популяций. То есть это не в неволе, это многолетние наблюдения за дикими популяциями амфибий и рептилий. Это колоссальный труд: каждую лягушечку нужно пометить, а потом год за годом следить, сколько из помеченных лягушек живы, сколько умерли, и таким образом отслеживать динамику смертности, рисовать кривую выживания и определять скорость старения.
Таких данных набралось много, всё это было обсчитано самой современной статистикой. Авторы искали связь продолжительности жизни и скорости старения с разными особенностями образа жизни, морфологии и биологии этих организмов. На рис. 3 в компактном виде представлены собранные данные2.
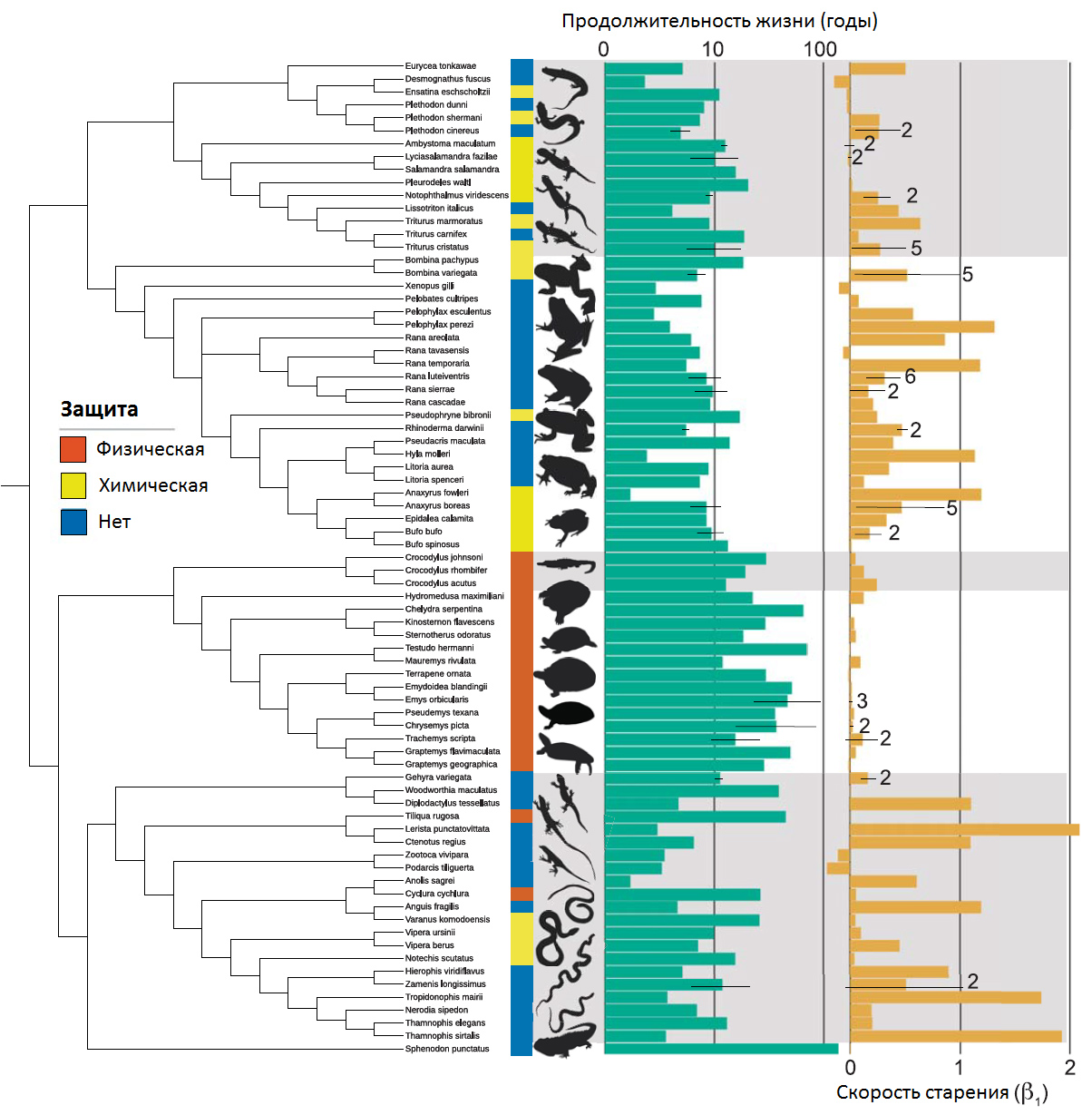
крокодилы, черепахи, чешуйчатые (ящерицы и змеи), клювоголовые (гаттерия). Из статьи в Science1
Зеленые столбики — максимальная продолжительность жизни каждого вида. Средняя слишком сильно зависит от условий, а максимальная — это видовая характеристика, более информативный показатель. Желтые столбики — скорость старения. Я говорил, что у людей смертность удваивается каждые семь лет, а у каких-нибудь лягушечек она может удваиваться каждые два года, например, — вот это и есть скорость старения.
Главный результат состоит том, что виды с хорошей защитой (имеющие прочную чешую, мощный панцирь, как у черепах, или ядовитые железы — т. е. хорошо защищенные, которых не так-то просто сожрать хищнику) при прочих равных живут дольше и стареют медленнее, чем незащищенные, которых легче съесть.
И вот что еще интересно. Существуют еще физиологические, метаболические гипотезы старения. То, что я изложил по поводу причин старения, — это взгляд эволюциониста (почему отбор не справляется со старением). А если вы спросите у физиолога, у биохимика, у клеточного биолога, каковы причины старения, он вам расскажет совсем другую историю. Он начнет говорить про накопление поврежденных молекул, про соматические мутации, повреждения ДНК, про митохондрии, производящие свободные радикалы, которые всё окисляют, про окислительный стресс и т. д. Из этого физиологического взгляда следует со всей очевидностью, что, например, теплокровные должны стареть быстрее холоднокровных, потому что у теплокровных быстрее метаболизм, активнее клеточное дыхание, соответственно, митохондрии производят больше активных форм кислорода, должны быстрее накапливаться повреждения. То есть теплокровные должны стареть быстрее. И вот это не подтвердилось, когда хорошие данные по холоднокровным сравнили с собранными ранее данными по птицам и млекопитающим. Также не подтвердилось соображение, что холоднокровные из холодных районов стареют медленнее, чем холоднокровные из теплых районов. Хотя, казалось бы, должны, потому что при высокой температуре быстрее метаболизм. Получается очень интересно: интенсивность метаболизма, ведущая к ускоренному накоплению молекулярных повреждений, менее значимый фактор эволюции старения, чем сила отбора на способность с этими повреждениями справляться. Эволюционная теория старения Медавара — Уильямса работает, а физиологическая — как-то не очень. Я бы резюмировал всю эту историю таким образом: эволюция сильнее физиологии. То есть будь у вас даже теплокровность, высокий метаболизм, куча свободных радикалов, но если есть достаточно сильный отбор на замедление старения — у вас будет замедленное старение. Эволюция найдет способ побороться с этими радикалами, повреждениями, с этими укорачивающимися теломерами — и что там еще у нас накапливается в процессе старения. А если отбор слабый, то даже при медленном метаболизме организм будет стареть быстро.
Вторая работа3 на эту же тему вышла уже в 2023 году. Она про млекопитающих. Опять же была собрана огромная база данных по млекопитающим — 974 вида. Здесь проверялась идея о том, что социальный (общественный) образ жизни способствует эволюции долголетия. Почему социальность должна способствовать эволюции долголетия? По той же самой причине: социальность — это очень хорошая защита. Общественный образ жизни защищает животное прежде всего от хищников. Защищает множеством способов — от простых до сильно продвинутых. Жизнь в обществе — это хорошая защита от хищников и для стадных копытных, у которых достаточно простые взаимоотношения в группе, и у приматов, у которых очень сложные взаимоотношения в группе. Поэтому мы ожидаем (исходя из классической эволюционной теории старения), что социальные виды будут стареть медленнее — в среднем и при прочих равных. В целом это подтвердилось (со всеми необходимыми поправками): социальные млекопитающие стареют медленнее и живут дольше, чем одиночки.
Здесь можно предположить, что это не социальность способствует эволюции долголетия, а наоборот, долголетие способствует эволюции социальности: животные сначала становятся долгоживущими, и это каким-то образом создает благоприятные условия для того, чтобы они стали после этого социальными. Это удалось проверить благодаря тому, что у авторов было эволюционное дерево, и они могли посмотреть, сколько было эволюционных переходов из долгоживущего в короткоживущее состояние и обратно, и из социального состояния в состояние одиночек и обратно. Оказалось, что переходы от короткоживущего состояния к долгоживущему чаще происходят у социальных, чем у одиночек. А обратная идея, что долголетие способствует эволюции социальности, не подтвердилась, потому что короткоживущие одиночки и долгоживущие одиночки с одинаковой частотой становятся социальными и наоборот.
Кстати, стимулом для эволюции долголетия может быть не только защищенность. У самых высокоорганизованных социальных млекопитающих, таких как люди и китообразные, отбор может поддерживать долголетие, потому что для группы очень выгодно, чтобы в ней были старые опытные особи, которые делятся своей мудростью или просто приводят группу к источнику во время засухи, потому что помнят, как спасались в прошлую засуху. То есть в этом случае уже работают факторы, связанные с обучением и опытом.
На заре эукариотной жизни
Следующая новость прошедшего года касается происхождения эукариот.
Их появление было величайшим событием эволюции земной жизни и в то же время одним из самых загадочных — хотя в значительной степени эта загадка уже разгадана. Но это, тем не менее, очень интересная проблема для эволюционных биологов: как появилась эукариотическая клетка — сложная клетка с ядром, из которой сделаны все сложные организмы.
У прокариотов, бактерий и архей клетка организована принципиально гораздо проще, и вроде бы существует пропасть между прокариотической и эукариотической клетками. Некоторое время назад она казалась настолько большой, что некоторые ведущие эволюционисты считали, что происхождение эукариот — это редчайшее явление, которое могло произойти только в одной из миллиардов миллиардов вселенных. То есть привлекалась некая случайность, потому что не было хороших переходных форм. Однако несколько лет назад произошел прорыв в этой области, потому что была открыта ранее совершенно неизвестная большая группа архей, или архебактерий, как их раньше называли, которые получили название асгардархеи. Первого представителя этой группы нашли в глубоководных отложениях около гидротермального источника на Срединно-Атлантическом хребте. В Северной Атлантике, уже практически в Арктике, есть такое гидротермальное поле, которое называется Замок Локи (по имени скандинавского бога), поэтому обнаруженных там архей назвали локиархеи. Потом нашли их родственников в других биотопах и стали их называть всех в честь скандинавских богов.
Это оказалась огромная разнообразная группа, доселе неизвестная. Первое время эти археи были известны только по геномам. То есть их обнаружили с помощью анализа проб грунта: из грунта выделяли всю ДНК, которая там есть, секвенировали, и из отсеквенированных кусочков собирали геномы этих доселе неведомых архей. В микроскоп их никто не видел, пока наконец японские исследователи после нескольких лет кропотливого труда не сумели вырастить одну асгардархею в лаборатории, рассмотрев ее затем в микроскоп. Это оказались довольно странные твари.
Уникальны они тем, что в их геномах очень много генов, которые до открытия асгардархей считались чисто эукариотическими. Почему и говорят, что асгардархеи — это переходные формы между прокариотами и эукариотами: у них много генов, которые неизвестны у других прокариот, но имеются у эукариот. В том числе там много генов, связанных с клеточным скелетом. У эукариот развит цитоскелет, состоящий из микрофиламентов и микротрубочек, он придает клетке подвижность, способность образовывать всякие выросты типа ложноножек, и он очень важен для эукариотической клетки.
Кроме того, в геноме асгардархей были обнаружены гены, связанные с разными мембранными пузырьками. У эукариот они обслуживают формирование разных мембранных структур, и поэтому думали, глядя только на геномы, что, может быть, у этих асгардархей, чем черт не шутит, даже ядро клеточное есть или какие-то мембранные органеллы. Оказалось, что ядра никакого нет, мембранных структур внутри клетки нет, но зато есть длинные, иногда даже ветвящиеся цитоплазматически выросты, подобные щупальцам. Но разглядеть это всё до сих пор не могли.
В 2022 году вышла статья4 про второй вид асгардархей, который тоже сумели вырастить в лаборатории. Культура этого второго вида лучше, численность клеток больше, и можно было более детально исследовать эти клетки. Удалось показать, что эти клеточные выросты, которые есть у обеих асгардархей, выращенных в лаборатории, окружены мембранами, что внутри них цитоплазма с рибосомами. Более того, оказалось, что клетки асгардархей этими выростами могут сливаться, т. е. образовывать цитоплазматические мостики между клетками. Внутри этих выростов есть микрофиламенты, состоящие из актина — белка, который играет важнейшую роль в построении цитоскелета. То есть эти сложные ветвящиеся выросты асгардархей поддерживаются настоящим динамическим цитоскелетом. Эти филаменты, по-видимому, могут удлиняться и сокращаться.
Возможно, эти длинные ветвящиеся выросты и цитоскелет, похожий на эукариотический, были главными эволюционными ноу-хау этой группы архей с самого начала ее существования. За счет этих выростов асгардархеи, по-видимому, могут налаживать сложные тонкие взаимоотношения с окружающей средой, друг с другом и с другими микроорганизмами. Ведь они живут в симбиозе с разными другими микробами — в одиночку либо вообще не растут, либо растут очень плохо. У них очень тесные трофические взаимоотношения с другими организмами в этих странных местах обитания, и эти щупальца помогают им налаживать взаимодействия с окружающей средой.
Новые данные хорошо согласуются с одной из старых гипотез о происхождении эукариот. Вообще, гипотез о происхождении эукариот опубликовано дикое количество, может быть, даже больше, чем гипотез о причинах массовых вымираний. Но одна из моделей, довольно известная, состояла в том, что предки эукариот приобрели митохондрии не путем фагоцитоза, не путем заглатывания бактерии, которая стала симбионтом, а по-другому. У клетки археи были выросты, и этими выростами она постепенно оплетала своего симбионта, бактерию, которая потом стала митохондрией. Затем эти выросты слились. Центральная часть архейной клетки стала ядром, остатки просветов между слившимися щупальцами стали эндоплазматической сетью — и вот так появилась эукариотическая клетка.
Эта гипотеза была достаточно популярна, но оставалась умозрительной. Когда увидели, что прокариоты, которые ближе всех к эукариотам и, по-видимому, похожи на предков эукариот, имеют длинные ветвящиеся, сливающиеся щупальца, это гипотеза заиграла новыми красками. Сейчас ее детализируют и развивают, гипотеза становится всё больше похожей на правду.
А почему важно, что там цитоплазматические мостики? Дело в том, что одно из уникальных свойств эукариот — это то, что называется половым размножением. Эукариотический секс — совершенно особый, чисто эукариотический способ перетасовки наследственного материала. У прокариот есть гораздо более простые его аналоги. В поисках переходных форм между прокариотическим примитивным сексом (горизонтальным переносом генов) и эукариотическим сексом исследователи уже довольно давно обратили внимание на некоторых архей из группы галоархей. У них был обнаружен необычный способ обмена генетическим материалом с помощью плазматических мостиков, по которым могут передаваться большие фрагменты генома, а возможно, даже целые хромосомы. Эти галоархеи — полиплоиды, как мы сейчас знаем, у них в каждой клетке в среднем 17 копий генома. Каждая копия генома — это одна кольцевая хромосома, и они обмениваются через плазматические мостики кусками хромосом, может быть, даже целыми хромосомами. Были выдвинуты гипотезы, в том числе мной с коллегами, о том, как именно, через какие промежуточные этапы мог появиться эукариотический секс. Примерно из такого полового процесса, как у галоархеи Haloferax. Всё в этих гипотезах хорошо и красиво, но из них следовало проверяемое следствие: у асгардархей, которые по отношению к эукариотам примерно как шимпанзе по отношению к людям, — не предки, но формы, происходящие от общего предка (тоже должны быть цитоплазматические мостики, чтобы у них был секс, похожий на эукариотический). И вот, пожалуйста, нашли подтверждение.
Эволюция за четыре дня
Следующая работа5 — это очень занятный эволюционный эксперимент. Пожалуй, я не буду вдаваться в детали, потому что они, может быть, не так интересны далеким от этих тем людям, но вот что меня привлекло в этой работе.
Это эволюционный эксперимент, который продолжался четыре дня, — вот как далеко мы ушли от идеи о том, что эволюция — это очень-очень медленный процесс, который заметен только в масштабе миллионов лет. Оказывается, бывают настолько быстро эволюционирующие организмы — особенно когда речь идет о эволюционной «гонке вооружений». В данном случае изучалась эволюционная «гонка вооружений» между бактериями и фагами, которые на этих бактерий нападают и в них размножаются, убивая их. Бактерии вырабатывают защиту от фагов с помощью своей замечательной системы CRISPR (clustered regularly interspaced short palindromic repeats). Это система приобретенного наследуемого иммунитета у бактерий. Я не буду рассказывать, что такое система CRISPR, чтобы сталось время на другое, это легко найти. В этом эксперименте изучалась коэволюция фагов с бактериями, имеющими систему CRISPR. За четыре дня эволюции там сменяется достаточно много поколений, и происходит куча интересных вещей, происходят эволюционные изменения и у бактерий, и у фагов. Авторы искали закономерности в этих процессах, подробнее можете прочитать соответствующие новости на сайте elementy.ru.
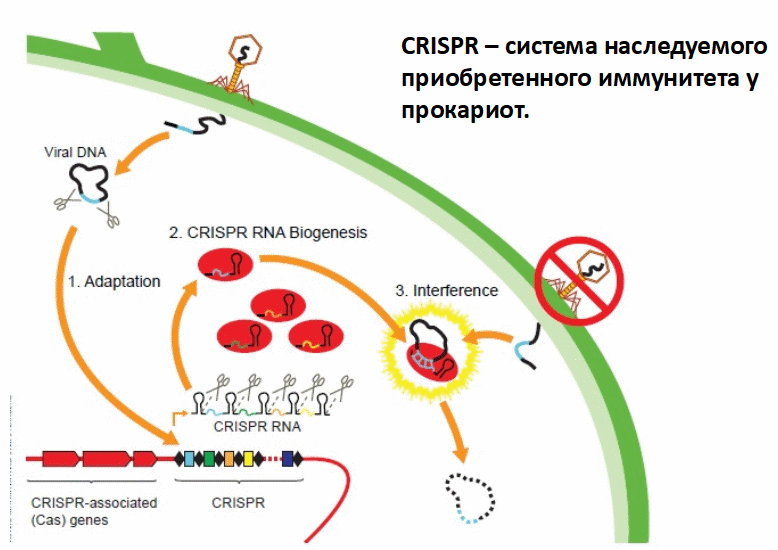
1) фрагменты вирусной ДНК вставляются в CRISPR-кластер («адаптация»);
2) CRISPR РНК синтезируются, процессируются, объединяются с белками («биогенез»);
3) Комплементарные участки вирусной ДНК атакуются и уничтожаются («интерференция»).
Илл. Asako Miyakawa and Sam Sternberg. sciencereview.berkeley.edu/read/fall-2012/germ-warfare
Первые зачатки рук и ног
Следующий блок будет из нескольких палеонтологических исследований. Палеонтологи в 2022 году тоже не бездельничали и много интересного откопали. Одна любопытная находка6 сделана в Китае. Вообще китайская палеонтология сейчас очень быстро развивается, китайские палеонтологи всё время открывают что-то новое и удивительное. На этот раз они описали панцирную бесчелюстную рыбу.
Панцирные бесчелюстные рыбы, или остракодермы, — это большая и разнообразная группа примитивных позвоночных, у них еще не было челюстей. От этих бесчелюстных до наших дней дожили миноги и миксины — они тоже позвоночные, но еще не имеют челюстей.
У панцирных бесчелюстных имелся костяной панцирь, покрывающий либо переднюю часть тела, либо почти всё тело. Это была большая разнообразная группа, от которой потом произошли и мы с вами. Новая панцирная бесчелюстная рыба из раннего силура Китая интересна тем, что она очень хорошей сохранности. На рис. 6 — изображение одного из экземпляров, вид с брюшной стороны.
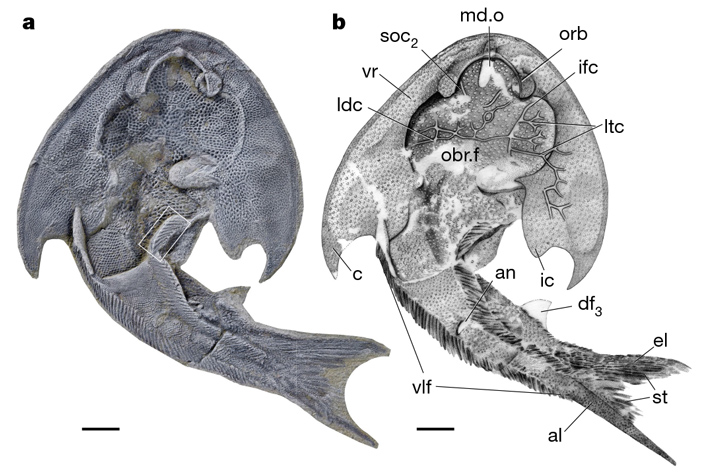
Самая интересная морфологическая особенность у нее — вдоль брюшка. У нее есть головной щит, парных плавников нет, но вдоль брюха идут две длинные плавниковые складки — два длинных плавника, которые тянутся от головного щита до хвоста.
Такая морфология раньше в таких деталях ни у кого не была описана. Она хорошо согласуется со старой доброй теорией происхождения парных конечностей позвоночных. В старых учебниках уже давно рисовали эту схему: как могли появиться парные конечности — передние и задние лапы. Они произошли от грудных и брюшных плавников челюстноротых рыб, а гипотеза состояла в том, что был предок, у которого было две плавниковые складки по бокам тела, которые потом разделились, компактизовались и превратились в грудные и брюшные плавники. В пользу этой схемы свидетельствовала интересная группа ископаемых рыб — акантоды. Это уже челюстноротые рыбы, вымершая группа, у них помимо парных грудных и парных брюшных плавников на линии, соединяющей парные плавники, было еще несколько также парных плавников в виде шипов. У этих рыб все плавники — в виде таких подвижных колючек, как будто действительно была единая плавниковая складка, которая потом поделилась на несколько отдельных парных плавников. В дальнейшей эволюции промежуточные плавники редуцировались, а самые передние и самые задние остались и стали в конце концов нашими руками и ногами. Морфология этого описанного тут туцзяасписа7, как его назвали, согласуется с этой идеей.
А еще известны Osteostraci, это другая группа остракодерм — самая продвинутая, — по-видимому, та, от которой, собственно, и произошли челюстноротые рыбы. У Osteostraci уже есть парные грудные плавники, но еще нет парных брюшных. Есть парные конечности, но только пара «рук»; «ног» еще нет. И вот сейчас, уже зная морфологию туцзяасписа, обратили внимание, что у этих рыб тоже есть нечто похожее на плавниковую складку, которая тянется вдоль брюха позади уже оформившихся грудных плавников. Это пример того, как новые факты подтверждают красивые старые идеи. Люблю такие случаи.
Александр Марков, докт. биол. наук
Окончание — в следующем номере
1 Reinke B. A., Cayuela H., Janzen F. J. et al. Diverse aging rates in ectothermic tetrapods provide insights for the evolution of aging and longevity // Science. 2022.
2 elementy.ru/novosti_nauki/433985/Pantsiri_i_yady_sposobstvuyut_evolyutsii_dolgoletiya_u_amfibiy_i_reptiliy/t5272064/Sergey_Kolenov
3 Zhu P. et al. Correlated evolution of social organization and lifespan in mammals // Nature Communications. 2023.
4 Rodrigues-Oliveira T. et al. Actin cytoskeleton and complex cell architecture in an Asgard archaeon // Nature. 2022.
5 Guillemet M. et al. Competition and coevolution drive the evolution and the diversification of CRISPR immunity. 2022.
6 Gai Z. et al. Galeaspid anatomy and the origin of vertebrate paired appendages // Nature. 2022
7 elementy.ru/novosti_nauki/434025/Pantsirnoe_beschelyustnoe_tutszyaaspis_prolivaet_svet_na_proiskhozhdenie_konechnostey_pozvonochnykh

 (7 оценок, среднее: 4,29 из 5)
(7 оценок, среднее: 4,29 из 5)
Думаю, для многих будет сложно разобраться по тексту в таких понятиях как эволюция, адаптация, отбор а пр. А вообще – очень интересный обзор действительно очень интересных работ.
В контексте имеющейся дискуссии могу отметить, что данный обзор хорошо показывает, как много мы еще не знаем. Вот, например, вопросы старения. Про отбор все хорошо. А каковы генетические детерминанты старения-нестранения и сроков? В тексте ни слова. И не случайно.
Вряд ли сюда может попасть кто-то не понимающий слов эволюция, адаптация и отбор, разве что они употреблены в каком-то неизвестном специальном смысле.
Разве то, что тут сказано про отбор, это не hand-waving в чистом виде? Можно ли вообще что-то доказательно утверждать про отбор? Бывают ли тут прямые, а не косвенные, доказательства; причинно-следственные связи, а не корреляции?
Читал про одну философскую историю, но, к сожалению, без подробностей. Известный Поппер предложил в своё время известный критерий фальсифицируемости (имея в виду марксизм-ленинизм). Однако ему доказали, что дарвинизм также не выдерживает такой проверки, после чего ему пришлось свой критерий значительно обессмыслить. Какая была аргументация, я не знаю.
Я, собственно, так и писал, что вся эволюционная наука есть hand-waving. Некоторые обобщения эмпирических данных, которые обычно допускают множество толкований. Отбор, впрочем, существует. Мы сами его осуществляем, выводя разные породы собак и лошадей.
Вот что такое “генетические детерминанты”, это я не знаю, а сказано так:
“Эволюция найдет способ побороться с этими радикалами, повреждениями, с этими укорачивающимися теломерами — и что там еще у нас накапливается в процессе старения.”
Видимо, автор имеет в виду, что принципиальная возможность как старения, так и борьбы организма с ним – ясна, а какие механизмы (может быть, и неизвестные) имеют место в каждом отдельном случае – это технические детали. Я так понял.
Заметка очень понравилась, – будит разум, – такие редкость.
Не исключено, старение – это не свойство и не признак, – это обязательная, неизбежная фаза или стадия жизни любых дискретностей, воспринимаемых как системы.
Например, старение каждого из нас – это как старение любой популяции, любого биоценоза, – ведь каждый из нас – это вселенная из 100 триллионов клеток, изо всех сил сохраняющая свой гомеостаз, – стремящаяся как можно дольше жить.
Замечу, мир почему-то бесконечно дискретен, квантован во всём, – и в пространстве и во времени, – похоже, именно поэтому мы воспринимаем его как причинно-следственный и изобрели числа, счет.
При желании, нетрудно растянуть свою индивидуальную жизнь на срок, превышающий среднюю продолжительность жизни, скажем в полтора раза, – достаточно перенастроить свой внутренний терморегулятор на гомеостатическую температуру с 36.6 С на 32.9 С. Другими словами, научится управлять своей теплокровностью в соответствии с правилом Вант-Гоффа с температурным коэффициентом равным 3.
Есть взаимодействие – есть старение. Нет взаимодействия – нет ничего, голая математическая абстракция. Всё стареет, если наблюдаемо, а стало быть находится во взаимодействии с хотя бы наблюдателем. Последний, к сожалению, тоже стареет ))
Хотя, с другой стороны, без взаимодействия его бы и не было. ИМХО (?)
Спасибо.
Очень познавательно и интересно.
Жизнь так многообразна и… Коротка!
Удачи Вам и здоровья!
просто процент умирающих каждый год будет одинаков, и численность когорты убывает экспоненциально
??
Очевидно, экспонента от отрицательного аргумента.