
Во времена, когда я и мои сверстники поступали на биологический факультет МГУ, каждому абитуриенту нужно было решить, что он хочет: бродить по лесам, горам и пустыням и бороздить моря в наблюдениях за живой природой или сидеть в лаборатории, капая из пипетки в пробирки, в поисках сути этой живой природы на уровне молекул. Надо было выбирать, куда поступать, — на первое, физиолого-биохимическое, или на второе, «полевое» отделение. Потом отделения объединили. Граница между полевой и лабораторной биологией в течение двух с половиной последних десятилетий стремительно размывается. Зоологам и ботаникам приходится осваивать молекулярные методы, чтобы получить ответы на вопросы, на которые не может ответить исследование морфологии и поведения живых объектов. Биология становится единой и гораздо более сложной, чем раньше, наукой о жизни.
Хорошо помню, как еще в 1990-е годы многие биологи, привыкшие к классической систематике животных, сопротивлялись нововведениям: «Ну была же такая стройная система, пришли молекулярные генетики — и всё испортили!» Классическая систематика (что в зоологии, что в ботанике) брала за основу морфологические признаки, и на основании их сходства в тех или иных группах делала вывод об их родстве. Классические филогенетические деревья строились на основе морфологии. Но с развитием технологий анализа ДНК появилась возможность сравнивать геномы — сначала по отдельным маркерам митохондриальной или ядерной ДНК, затем — по полностью секвенированным последовательностям. Сначала лишь для некоторых видов животных и растений, но со временем число полностью прочитанных геномов стремительно нарастало.
И выяснилось, что генетическое родство организмов друг с другом далеко не всегда очевидно, а казавшееся очевидным родство по внешнему сходству зачастую оказывается иллюзорным. Хотя, действительно, трудно воспринимается теперь уже широко растиражированное открытие, что ближайшими родственниками китов являются гиппопотамы и другие парнокопытные, так что по молекулярной филогенетике китообразных как отдельной группы теперь не существует, зато есть отряд китопарнокопытные (Cetartiodactyla)1. Но с генами не поспоришь. Филогения ставит задачу реконструировать ход эволюции, а эволюция оперирует генами. Следовательно, молекулярная филогения априори оказывается более объективной, чем морфологическая. И сегодня уже редкая статья в зоологическом или ботаническом журнале обходится без подтверждения результатов молекулярными данными.
О том, как эта молекулярно-генетическая революция воспринимается изнутри классической биологии, что изменилось, какие вещи пришлось пересмотреть и какие новые возможности открывают молекулярные методы, я поговорила с несколькими биологами, работающими в разных областях науки. Мы начали общаться на эту тему на конференции Беломорской биологической станции (ББС) МГУ, проходившей в сентябре 2023 года, в год 85-летнего юбилея биостанции 2. Среди участников конференции было много молодых ученых, которые уверенно и увлеченно рассказывали об исследованиях своих объектов на всех уровнях — от популяций и отдельных организмов до молекулярной генетики. Современная морская биология, как и биология вообще, превратилась в науку очень широкого профиля.

«Дело морфологов — уже не пытаться построить свою филогению по морфологии, а понять, как она могла получиться»

Александр Борисович Цетлин, докт. биол. наук, директор Беломорской биологической станции МГУ3, полагает, что благодаря молекулярным методам классические зоологи получили фантастический инструмент. И теперь дело морфологов — уже не пытаться построить свою отдельную филогению по морфологии, а пытаться понять, как именно могли возникнуть животные с определенным набором признаков.
— По вашей оценке, изменения, которые принесли молекулярно-генетические методы в зоологию, это революция?
— Да, действительно, революция. Я считаю, что классические зоологи получили в свои руки инструменты, о которых они даже не мечтали. И теперь с наслаждением ими пользуются. Я помню время, когда единственным молекулярным инструментом для оценки филогенетических связей был процент гибридизации ДНК. Помню, что я участвовал в такой работе: нужно было собрать один грамм трихоплаксов — фантастическое количество этих ничтожных существ, — чтобы добыть столько ДНК, чтобы можно было произвести гибридизацию. Это было в конце 1970-х — начале 1980-х. Сейчас этот инструмент стал абсолютно необходимым не только для того, чтобы выяснять филогению больших таксонов, но и для того, чтобы заниматься филогеграфией и филогеномикой, чтобы выяснять, кто, откуда и как расселялся. Это фантастический инструмент.
Кроме того, это инструмент систематики, который позволяет определять, что мы имеем дело с криптическими видами, видами-двойниками, которые мы не можем различить морфологически. И, наконец, это инструмент для решения вопросов о жизненных циклах. В некоторых группах беспозвоночных описаны 300–400 разных личинок — и втрое меньше взрослых организмов. И мы понятия не имеем, как эти личинки соотносятся со взрослыми животными. Личинок мы часто ловим в океаническом планктоне, который течение переносит на тысячи километров, а где живут взрослые животные, мы толком не знаем. Теперь в наших руках есть инструмент для решения этих вопросов. Кроме того, речь идет о выяснении жизненных циклов паразитов, потому что только методами метагеномики можно определить, обитает ли тот или иной вид паразитов в таком-то многоклеточном животном.
Тем не менее есть люди, которые пытаются этому оппонировать. Есть такой очень интересный философ от зоологии Кирк Фицью (Kirk Fitzhugh), блестящий знаток некоторых групп многощетинковых червей, и он сформировался как ученый в то время, когда в зоологию очень прочно вошли представления о необходимости использования формальной логики для решения всех научных вопросов. Применительно к зоологии, сравнительной анатомии и филогении живого мира это были методы кладистики. Это отдельная история, потому что попыток использовать формальную логику для решения вопросов филогении было несколько, и они были параллельны, в разных странах ученые одновременно к этому приходили.
— В чем именно применительно к зоологии заключается формальная логика?
— Формальная логика заключается в том, что научная работа состоит из формулирования гипотезы и ее проверки.
— Это было не очевидно?
— Это было абсолютно не очевидно. Идеи прогрессивной эволюции живого мира совершенно захватили ученый мир в последней трети XIX века. Вопросы к ним появились в тот момент, когда в науку вошло огромное количество новых фактов. В конце XIX века стали развиваться методы микротехники, и сведения о строении животных быстро пополнялись данными нового уровня и нового объема. К 1910–1920-м годам этих данных накопилось очень много. Огромный объем информации вошел в зоологию, ученые его переварили и стали анализировать. И почти одновременно в разных странах появляются люди, которые заметили: когда мы смотрим на это необозримое количество фактов, то наши привычные гипотезы очень плохо подтверждаются. Следовательно, наши гипотезы основаны на логических конструкциях, которые имеют изъяны. И мы должны отделить факты от наших фантазий и домыслов и вернуться к формальной логике. Так делают несколько классических немецких зоологов, а в России так делает Беклемишев. И дальше — эту историю я очень люблю — Беклемишев критикует «сказочную» сравнительную морфологию, в своей книге «Методология систематики» в 1935 году он от нее не оставляет камня на камне. А в 1948 году он публикует «Сравнительную анатомию беспозвоночных». И интересно, что эта его новая анатомия полна таких же недоказуемых утверждений и таких же прыжков логики, как и все конструкции предыдущих зоологов, которые он критиковал. Объяснить это просто — пока университет во время войны был в эвакуации, Беклемишев, будучи заведующим кафедрой зоологии беспозвоночных, читал студентам лекции по сравнительной анатомии. И это очень хорошо показывает, что «сказочная» зоология с красивыми филогенетическими конструкциями — это, в сущности, дидактический прием, который облегчает студентам освоение большого разнообразия фактов.
Но Кирк Фецью воспринял кладистику как очень хороший инструмент для науки. И когда появилась молекулярная биология и систематики стали ею пользоваться, ему показалось, что прежняя красивая хрустальной чистоты конструкция беспардонно ломается фактами, которые очень трудно трактовать с точки зрения формальной логики.
— Молекулярные методы разрушили эту красивую конструкцию?
— Они ее не разрушили, они ее, на мой взгляд, в значительной степени делают ненужной. Вот вы строите таблицу морфологических признаков, на их основании строите кладистические деревья. И вдруг оказывается, что вы их строили-строили, а на самом деле всё по-другому. Потому что молекулярная филогения — ей плевать на ваши признаки, и с ней очень сложно спорить.
— Но ведь какие-то вещи подтверждаются молекулярной филогенией?
— Очень часто подтверждаются. Но Кирка и его последователей это не устраивает. Он считает, что всё должно выглядеть так: вы должны высказать гипотезу, что, возможно, в вашем материале есть признаки нового таксона, вы должны сформулировать признаки, по которым вы будете испытывать эту гипотезу, и, наконец, вы должны прийти к выводу, что такие-то и такие-то процедуры, исполненные в вашей работе, показывают, что это новый таксон, который отличается от всех остальных тем-то и тем-то. Я до конца не понимаю, почему ему кажется, что молекулярная наука несовместима с этой работой, но у него много последователей. Очевидно, адепты этой философии считают статьи с молекулярными данными статьями из другого мира. Но в целом это всё же маргинальная часть ученых.
— Не изменилась ли концепция вида с появлением молекулярных методов?
— Это хороший вопрос. С концепцией вида вообще плохо. Каждый систематик старается не трогать этот больной вопрос, потому что у нас очень мало инструментов для его решения. Потому что живые существа чрезвычайно разные. То, что вам кажется абсолютно логичным применительно к беспозвоночным, неприменимо к другим: скрещиваются — один вид, не скрещиваются — разные, дальше всё размывается. Что там со скрещиванием у бактерий, понять трудно, а высшие млекопитающие отлично скрещиваются. Таких вопросов очень много, и мне кажется, все более-менее по умолчанию смирились, что надо в каждом конкретном случае приходить к соглашению, что мы будем в данной группе считать видом, и не пытаться экстраполировать полученное знание на весь остальной мир.
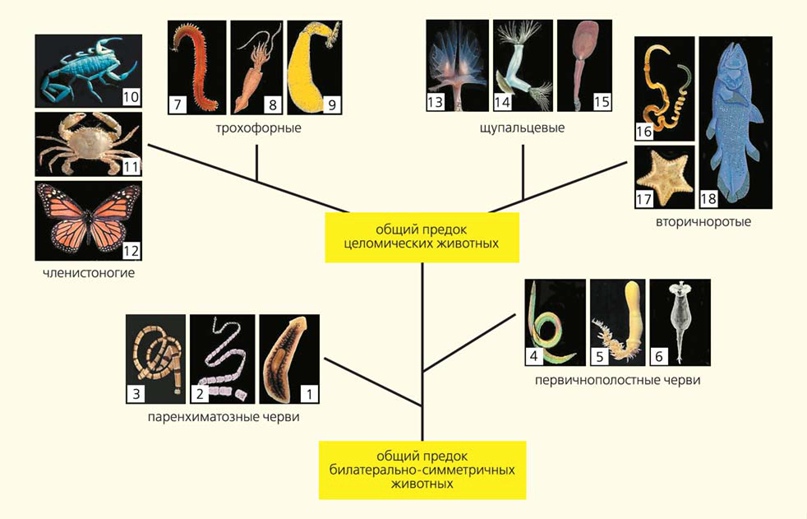
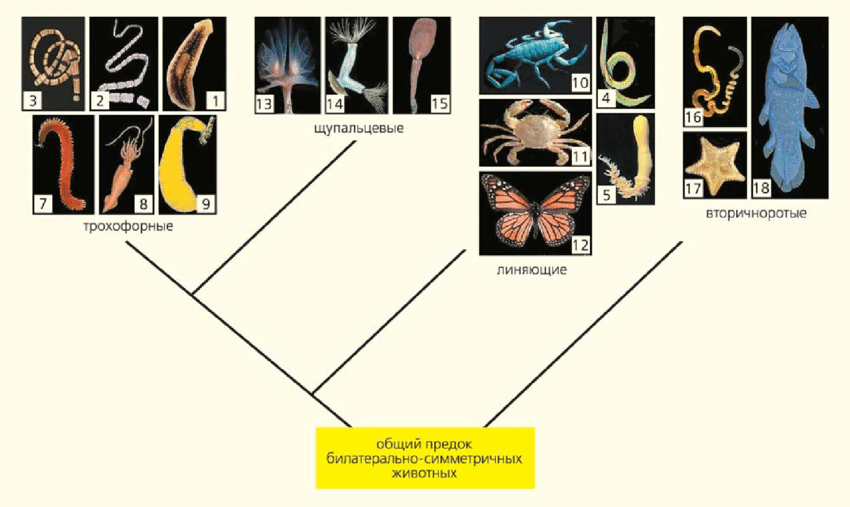
Классические (вверху) и новые (внизу) представления о системе и филогении билатерально-симметричных животных. 1 — свободноживущие плоские черви (турбеллярии), 2 — паразитические плоские черви, 3 — немертины, 4 — круглые черви (нематоды), 5 — головобрюхие черви, 6 — коловратки, 7 — кольчатые черви, 8 — моллюски, 9 — сипункулиды, 10 — хелицеровые, 11 — ракообразные, 12 — насекомые, 13 — форониды, 14 — мшанки, 15 — брахиоподы, 16 — полухордовые, 17 — иглокожие, 18 — хордовые. На схеме представлены далеко не все группы животных (Малахов В. В. Революция в зоологии: новая система билатерий // Природа. 2009. № 3. С. 40–54)
— А если посмотреть на крупные систематические таксоны, то что принципиально изменилось? Пятнадцать лет назад в журнале «Природа»4 Владимир Васильевич Малахов, завкафедрой беспозвоночных животных в МГУ, опубликовал две схемы филогении билатеральных животных — старую и новую, молекулярную. Как бы вы прокомментировали: какие принципиальные изменения появились в новой схеме?
— Владимир Васильевич в своей статье абсолютно всё правильно написал: эта старая схема, в сущности, созданная в конце XIX века, отображала общее представление о прогрессивной эволюции. Кроме того, были еще представления о консервативности тех или иных систем органов. Они были основаны на тех фактах, которые имелись в то время у ученых на руках. Казалось, что нервная система, расположенная на брюшной стороне и устроенная как брюшная нервная цепочка, — это некая фундаментальная характеристика, и все животные с таким типом нервной системы должны быть каким-то образом между собой связаны, что и отображает эта филогенетическая схема, где у нас находятся вместе черви сегментированные и моллюски — все трохофорные животные, у кого нервная система на животе и более-менее лестничного типа. Членистоногие оказались к ним самой близкой группой, потому что у них такой же тип нервной системы, и это казалось очень существенным признаком.
Фактически первый звонок о том, что этот вопрос требует дополнительного обсуждения и обдумывания, прозвенел в тот момент, когда были открыты гомеобоксные гены. Это гены, которые в процессе развития контролируют сегментацию, они отвечают за образование метамеров. Они были найдены во всех органах у всех животных, чуть ли не у простейших. И оказалось, что процесс метамеризации, аннуляции — это некая фундаментальная возможность, которая есть у любых живых существ — во всяком случае, у всех многоклеточных животных. Таким образом, вопрос о глубокой связи между артроподами (членистоногими) и аннелидами (кольчатыми червями) уже подвис, и, в сущности, надо было заново его обсуждать.
— Вы говорили, что их объединяли на основе сходства строения нервной системы, и получается, что главным общим признаком строения была сегментация, так?
— Всё вместе — сегментированное тело, т. е. состоящее из члеников, брюшная нервная цепочка, парные придатки на каждом сегменте. Но гомеобоксные гены показали, что это не бог весть что, а свойство, которое есть у всех многоклеточных животных. Дальше с развитием молекулярной филогенетики что произошло? Появилась группа «линяющие», в которую вошли членистоногие и оказались вместе с нематодами (круглыми червями).
— А нематоды линяют?
— Нематоды линяют. Но вопрос ведь не в том, что одни линяют, а другие не линяют. Например, медицинские пиявки (в отличие от других пиявок), которые кольчатые черви, линяют так же, как артроподы. Но они теперь в другой группе.
— Название «линяющие», получается, условное?
— Конечно. Фундаментальный факт состоит в том, насколько организмы отличаются по данным молекулярной генетики, а не в том, линяют они или не линяют. Сейчас ситуация такая. Мы полагаем, что данные молекулярной филогенетики, особенно если они получены по большому количеству генов или по целому геному, дают нам основание строить филогению. И получается, что дело морфологов — уже не пытаться построить свою отдельную филогению по морфологии, а пытаться понять, как эти животные с таким набором признаков могли получиться. Конечно, может быть, что через пятьдесят лет всё будет по-другому, но сейчас нам кажется, что не нужно спорить с данными молекулярных филогенетиков. Мы видим, что построенные по молекулярным данным деревья где-то уже устоялись, где-то они находятся в процессе уточнения. Большие ветви более-менее устоялись.
— Что касается больших ветвей: помимо появления новой группы «линяющие» и размежевания кольчатых червей с членистоногими, что еще принципиально поменялось?
— Скажем, было представление о том, что турбеллярии (ресничные плоские черви) — очень примитивная группа, и у самых примитивных еще нет кишечника, а потом из них получаются, во-первых, более сложные кишечные турбеллярии, а во-вторых — паразитические черви, у которых в связи с паразитическим образом жизни теряется кишечник. Это всё здорово, но еще в 1980-е годы было показано, что бескишечные турбеллярии очень трудно называть действительно примитивными животными, потому что у них есть очень специфический способ восстановления центральной паренхимы. У них есть некая центральная паренхима, в которую попадает еда, и там она переваривается. И оказалось, что в этой зоне вообще нет пролиферации клеток, а вся пролиферация происходит только в периферической паренхиме, близко к внешнему эпителию. Эти работы были проведены в Зоологическом институте в Санкт-Петербурге под руководством Юрия Викторовича Мамкаева очень тонкими и сложными методами, при помощи авторадиографии, они метили клетки. И оказалось, что турбеллярии — это очень специализированные животные с очень сложным механизмом восстановления клеточных популяций. И разговоры о том, что они какие-то совсем примитивные, как-то хромают, да? Получается, что они, скорее всего, вторично упрощенные животные.
— То есть какой ни возьми морфологический признак, который кажется очень фундаментальным, оказывается, что для филогении он не работает, так?
— Понимаете, пока у нас нет сомнений, что развитие идет в одном направлении, он работает. Как только мы начинаем задумываться, оказывается, что не всегда, казалось бы, очевидный факт двигает общее представление. Еще тридцать лет назад в школьных учебниках рисовали, что турбелляриии самые примитивные, хотя для тех ученых, которые этим вопросом занимаются, факты были уже другими.
— Схема молекулярной филогении, опубликованная Владимиром Васильевичем, сейчас принципиально не изменилась?
— Мне кажется, пока нет. Но это не значит, что она навечно такой останется. Трудно предположить, что будут катастрофические изменения, но мало ли что еще будет. Это всего лишь рабочая гипотеза, не больше.
«Если мы считаем, что с видом всё хорошо, значит, скорее всего, его еще толком не исследовали»

Анна Эльмировна Жадан, канд. биол. наук, ст. науч. сотр. Беломорской биологической станции МГУ.
— Какие изменения в вашей области науки произошли с появлением молекулярных методов?
— Для Annelida (кольчатых червей) за последние 20–25 лет молекулярные методы полностью перестроили общую систему. Нас когда-то учили, что кольчатые черви делятся на многощетинковых (полихет) и поясковых (клителлят), а они в свою очередь на малощетинковых (олигохет) и пиявок. А полихеты делились на эррантных (бродячих) и седентарных (сидячих). Но уже во второй половине XX века считали, что это так себе группировки, не очень хорошие, скорее экологические, чем таксономические. Кроме того, были группы ранга классов и типов неясного таксономического положения, более или менее родственные кольчатым червям, такие как сипункулиды, эхиуриды, сибоглиниды (это погонофоры, вестиментиферы, оседакс), мизостомиды. Теперь же стало понятно, что все эти группы входят в состав аннелид. Более того, таксоны эррантных и седентарных полихет снова вернули в систему, но уже с новым смыслом. И сейчас сложилась более-менее общепринятая система, что у аннелид есть группы базальной радиации, и есть эррантные и седентарные — группы, к которым принадлежит большинство видов. Интересно, что малощетинковые черви и пиявки вошли в состав группы седентарных. Получилось, что таксон Polychaeta (многощетинковые черви) потерял свой смысл, так как стал тождественен Annelida. И хотя это слово по старой традиции используется и всем понятно, в строгой научной терминологии мы не должны его употреблять.


Фото А.Э. Жадан, сделаны в Кувейте
На схеме внизу — современная система аннелид (по Weigert, Bleidorn 2016). В желтом прямоугольнике — группы базальной радиации. Седентарные (в синем) и эррантные (в зеленом) раскрыты по сторонам от общей схемы. Красным подчеркнуты таксоны, которые раньше были отдельными классами или типами. Получается, что полихеты — это все аннелиды, которые не принадлежат к этим группам, т. е. этот таксон не включает всех потомков одного предка. Такие таксоны называют парафилетическими и они не должны использоваться в систематике.


— То есть то, что было показано молекулярно-генетическими методами, стало общепринятым?
— Да, но для этого сделали их транскриптомику, не просто посмотрели ген цитохромоксидазы CO-1. И определили какие-то очень глубокие ветви, которые возникли до кембрия или в очень раннем кембрии. В разных группах аннелид всё очень по-разному. Есть семейство Onuphidae из отряда Eunicida, и про них опубликована статья под названием «Когда молекулярные методы подтверждают морфологию»5. Там реально нашли строгие соответствия — вот молекулярные деревья, и у каждой веточки находятся морфологические подтверждения. А есть семействo Orbiniidae из отряда Orbiniida, где всё наоборот. Там вообще нет хорошей системы внутри семейства, там нет нормальных подсемейств, нет нормальных родов, там были какие-то системы, но, во-первых, они все друг другу противоречат, а во-вторых, молекулярные данные ничего из этого не подтверждают. Сейчас ими занялись подробнее, на большем числе объектов, делают целиком митохондриальные геномы, ядерные геномы. Уже появилось дерево, хотя далеко не все таксоны там представлены, но в этом уже начинает проявляться какой-то смысл.
На уровне видов чаще всего получается так: если мы считаем, что с видом всё хорошо, значит, скорее всего, его еще толком не исследовали. Дело в том, что большинство видов было описано из европейских вод, а затем эти названия присваивали похожим червям из всех морей и океанов. Есть исключения, когда оказывается, что даже в Америке и в Европе живет один и тот же вид. Но чаще оказывается так: то, что именовали каким-то одним названием, распадается на комплексы криптических или псевдокриптических видов. Псевдо — это если у них есть маленькие различия, просто их раньше либо не находили, либо не обращали внимания. Вот один пример: полихеты рода Eteone. Мы считали, что у нас в Белом море живет один вид Eteone longa. А оказалось, что там пять видов, из которых три новых для науки, и с этим надо разбираться, разбираться и разбираться.
«Взаимная провокация хороша сама по себе»

Михаил Владимирович Калякин, докт. биол. наук, директор Зоологического музея МГУ, отвечал на мои вопросы практически на бегу, в перерыве между докладами на конференции; мы буквально лавировали между постерами и участниками мероприятия.
— Каков взгляд орнитолога на молекулярно-генетическую революцию?
— Возможность использовать молекулярно-генетические методы в зоологии приводит к тому, что появляется вторая система координат: одна по морфологии, другая по генам. Если говорить про птиц, то для них эта система третья, потому что орнитологи используют еще и вокализацию. Но даже если мы отбросим голоса и оставим две системы координат, то возникает довольно занятная ситуация. Если данные, полученные методами молекулярной генетики, не сходятся с морфологическими, генетики говорят: смотрите, мы показали, что это не так, и нужно всё пересмотреть. А если сходятся, говорят: видите, и морфология подтверждает наши выводы. И появляется бинарная система, при которой все счастливы, потому что всегда можно соотнестись друг с другом.
Время от времени, когда генетики получают новые данные, то морфологи сначала просто отмахиваются от них. Ну, например, когда генетики приходят к выводу, что кондоры Нового Света — это аисты. Орнитологи, которые тридцать лет думали по-другому, говорят: ну что за ерунда?! Но если покопаться в морфологии внимательно, то иногда оказывается, что генетики дают нам важный сигнал: ребята, вы базируетесь на данных, которые получили в конце XIX века и с тех пор не пересматривали. А вы поковыряйтесь чуть-чуть еще. Не только в зоологии, но и в ботанике так происходит, что приходится многое пересматривать, особенно если речь идет о форме, базальной для какой-то из филогенетических веток. Поэтому вот эта взаимная провокация — она хороша сама по себе.
Я смотрю на это со стороны как функциональный морфолог, мои исследования посвящены ротовому аппарату птиц. Мы работаем давно проверенным методом: видим структуру, видим косточки, связки и просто из геометрии понимаем, что этот мускул поднимает челюсть, этот опускает, этот управляет движениями языка. Довольно простые механические построения позволяют нам однозначно трактовать функцию какого-то объекта. Когда я этими же глазами смотрю на то, что делают генетики, я понимаю, что там гигантские молекулы, которые как-то должны взаимодействовать. И вот когда же мы поймем механику этих взаимодействий на молекулярном уровне и сможем таким путем понять функции этой структуры? Не знаю, сколько должно пройти времени и какой математический аппарат для этого должен быть создан. Мы и сейчас многое понимаем, но картина всё время уточняется, и еще долго будет уточняться. На этом уровне ясно, что у нас нет абсолютно стройной гипотезы «от молекул до того, как действует конкретный орган», а тем более — организм. Хотя какие-то трассы уже прочерчены, когда мы знаем, какой ген что регулирует, и это большое счастье. И тогда те признаки, которыми пользуются морфологи, начинают приобретать значимое генетическое обоснование.
Вот пример. Есть птицы из группы тимелий, обитающие в Юго-Восточной Азии, в Гималаях, их много, несколько десятков родов. Это очень разнообразная группа, они живут в горах, и на каждой горке может обитать свой набор. Всё очень хорошо для эволюционных построений. Так вот, в 1990-е годы на них набросились генетики и что-то понаделали, в основном используя митохондриальную ДНК. Прошло несколько лет, и выяснилось, что для птиц митохондриалку использовать плохо. Посмотрели несколько ядерных генов, всё немного перевернулось, и теперь мы знаем, что ситуация явно требует дальнейших уточнений, и надеемся на полногеномное секвенирование.




Человек, далекий от этой кухни, глядящий на нее с другого берега, иногда еще и в бинокль, просто должен спокойно ждать, когда ребята дойдут до ситуации, когда они сами свои предыдущие построения не будут пересматривать каждые десять лет. Когда изобретатель отточил свой инструмент окончательно, убедился, что он действительно работает, тогда мы его берем и применяем. Пока что на автозаводе ребята шуршат, раздаются какие-то крики, несут новую деталь, здесь надо сварить, это надо выкинуть… Когда автомобиль выедет с завода, а потом будет участвовать в гонках и не развалится, тогда я его куплю и в него сяду. А пока там какие-то детали высыпаются, ну, ждем.
— Как сегодня обстоит дело с концепцией вида?
— Могу сказать про область моих научных интересов. По птицам Юго-Восточной Азии каждый год в большом журнале публикуется обзор того, как поменялись названия многих групп, потому что генетики изучили какой-то вид, этот вид распался на два, а таксономия основана на почти юридических основаниях. Концепции вида это касается в том смысле, что есть биологическая, а есть филогенетическая, и вечный спор между ними, который никогда не разрешится. Вот, есть линия, которая генетически отделяется от другой. Но если мы здесь режем, тогда получается два вида, а если здесь — вид разделяется на подвиды. И что с этим делать, непонятно.
Например, есть близкие виды пеночек, между которыми вымерших форм, наверное, нет, потому что они и морфологически и генетически отличаются совсем чуть-чуть. Но при этом в один генетический кластер попадают привычные нам желто-зеленые, блеклые пеночки и две ярких, цветных. А в другой кластер попадают яркие, цветные, и среди них пара-тройка этих бледно-зеленых. Любой классический орнитолог скажет: ну, это ерунда, так не бывает. А генетики говорят: вот так они кластеризуются. Орнитолог на это возражает: «Ну вы как работаете с вашими деревьями? Там же всё на вероятности? Может быть, вы что-то подработали, не нравилась эта веточка, обернули ее на 90 градусов — и вот результат: птицы в одном кластере — живут по-разному, поют по-разному, и вообще…» Морфологам трудно это понять — у них или есть косточка, или ее нет. А вот бывает гибридизация, когда какие-то кусочки генома от одного вида переходят к другому. Может быть, вы попали как раз в такой ген, и у вас красная пеночка оказалась среди желтых, и наоборот? Вот когда сделают полногеномное исследование всех этих тридцати видов близких пеночек, не забывая об их географической изменчивости, когда разберутся, какой у них уровень различий — на 3%, на 7%, видовой он или не видовой, — тогда будет счастье и окончательное знание.
Надежда Маркина, канд. биол. наук
Продолжение следует
1 Гельфанд М. Молекулярная эволюция: как киты уходили под воду // Природа № 10, 2016. elementy.ru/nauchno-populyarnaya_biblioteka/434242/Molekulyarnaya_evolyutsiya_kak_kity_ukhodili_pod_vodu
3 См. также интервью Михаила Гельфанда с Александром Цетлиным в рамках проекта «Разговоры за жизнь»: trv-science.ru/2022/01/aleksandr-cetlin-razgovory-za-zhizn/
4 Малахов В. Революция в зоологии: Новая система билатерий // Природа, 2009, № 3. researchgate.net/publication/297735440_Revolucia_v_zoologii_Novaa_sistema_bilaterij
5 sciencedirect.com/science/article/pii/S1055790315003140

 (3 оценок, среднее: 4,33 из 5)
(3 оценок, среднее: 4,33 из 5)