Цикл интервью Надежды Маркиной о молекулярных методах в современной биологии подытоживает беседа с Михаилом Гельфандом, докт. биол. наук, вице-президентом по биомедицинским исследованиям Сколтеха, членом Европейской академии, почетным членом Международного общества вычислительной биологии. Михаилу Сергеевичу пришлось «за молекулярных генетиков» отвечать на некоторые вопросы, возникшие у предыдущих спикеров1.
— Все зоологи и ботаники, с которыми я говорила, признают, что молекулярные методы произвели революцию в филогении. Было сказано, что дело морфологов — не пытаться построить свою отдельную филогению по морфологии, а пытаться понять, как эта морфология могла получиться. Хотя были высказаны и некоторые «но». Первое: в молекулярных деревьях какие-то вещи быстро меняются, какие-то группы «прыгают» по дереву туда-сюда, и не следует ли подождать, пока всё устаканится, прежде чем этим деревьям доверять. И второе «но»: молекулярные деревья строятся на вероятностном подходе, а вероятность — это как бы не 100% уверенности. Что вы можете сказать на эти «но»?

— На первое «но» я могу ответить, как сиреневый иностранец в Торгсине: «Кароши люблю, плохой — нет». Плохие молекулярные филогении быстро перестраиваются, а хорошие молекулярные филогении не перестраиваются. Современные методы построения деревьев, помимо всего прочего, сообщают меру своей уверенности во внутренних узлах. Как раз то, что мы можем оценить надежность реконструкции, — это большое достоинство молекулярных деревьев. С морфологическими деревьями может доходить до мордобоя, но количественной оценки нет.

Второе. Любая новая таксономия, основанная на молекулярных деревьях, некоторое время активно обсуждается. Ну вот, гипотеза, что нематоды и членистоногие относятся к одной группе. Несколько лет люди ругались, рисовали разные деревья, потом это устаканилось, и, по-моему, сейчас уже стало общим местом. Но общим местом это стало у нас на глазах.
Есть разные гигиенические правила при построении молекулярных деревьев. Их надо проверять на устойчивость. Надо проверять, что они не зависят от того, каких представителей таксонов вы взяли. Бутстрепы делают, чтобы их оценивать. Но если правила гигиены соблюдать, то всё хорошо. А с морфологическими деревьями проблема вот какая. Удивительным образом оказывается, что какое бы молекулярное дерево ни было построено, классические зоологии и ботаники найдут морфологические признаки, которые будут этому дереву замечательным образом соответствовать. И тем самым видно, что деревья, основанные на морфологии, не безумно хороши.
Дальше: оказалось, что молекулярные деревья действительно ведут к пересмотру каких-то очень глубоких представлений на уровне типов. И, если я правильно понимаю, это более-менее устаканилось. А с другой стороны, они позволяют делать какие-то вещи, которые морфологи не могли.
Есть две моих любимых истории. Первая: киты оказались ближайшими родственниками бегемотов 2. Сейчас это уже совсем общее место, но история все-таки чудесная. А второе — змеи оказались ящерицами. И я подозреваю, что на самом деле есть много других таких же историй, про которые мы просто не знаем, они менее раскручены. Хотя это ведь позвоночные, а киты вообще млекопитающие. То есть речь о совсем недавних событиях. А с другой стороны, появились какие-то таксоны, которые люди просто не выделяли. Был кустик какой-то странный, амборелла, по внешним признакам его пихали то туда, то сюда, а оказалось, что эта самая ранняя ветка цветковых 3.

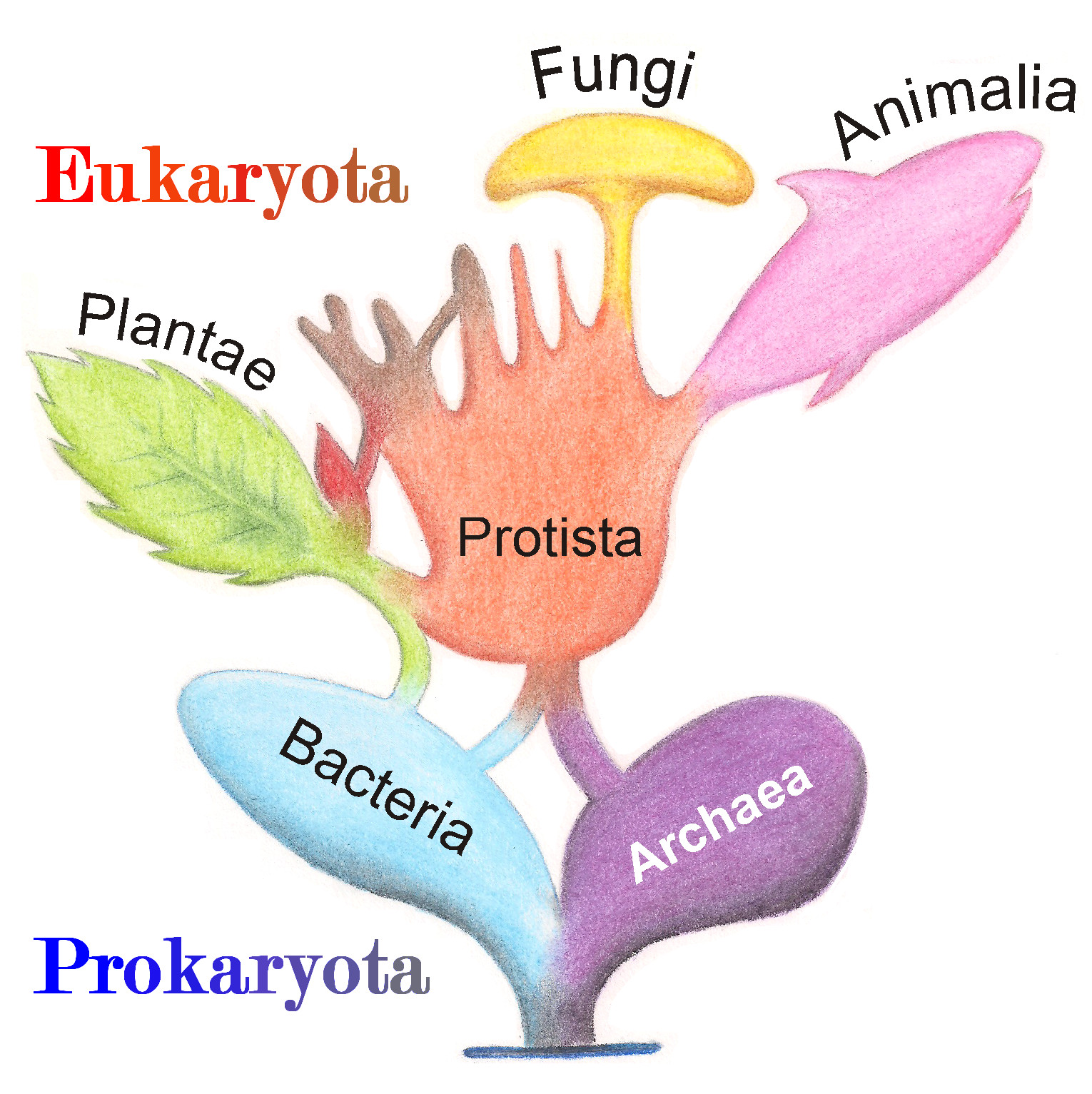
И это всё мы говорим про многоклеточных, у которых есть хоть какая-то морфология. А самое-то чудесное произошло не с ними, а с одноклеточными. Во-первых, простейшие, которые фактически были такой же помойкой, как линнеевские «черви», туда просто скидывали всё, что непонятно. Оказалось, что простейших очень много разных; открыли и открывают много новых типов. Взаимоотношения между ними не до конца понятны, но глубина разделения очевидна.
Потом еще одна моя любимая байка: грибы перестали быть низшими растениями и вообще перестали быть растениями, оказались ближе к животным. Стало ясно, что многоклеточность много раз возникала, чего никакие морфологи, по-видимому, в виду не имели: классическая картина была такой, что сначала появились многоклеточные, а потом они разделились на растения и животных. Но посчитали и показали, что всё не так. Вообще, это мировоззренческого уровня вещь. И это еще даже ничего, а вот где совсем всё стало по-другому — так это прокариоты.
— Да, там же некоторые таксоны вообще только молекулярно описаны.
— Там все крупные таксоны и взаимоотношения между ними описаны только молекулярно. А когда начали делать метагеномы, оказалось, что мы для большей части разнообразия просто не подозревали, что такое бывает. Если посмотреть на таксономию бактерий, то глубокие узлы там разрешаются плохо; очень глубокая реконструкция будет ненадежна. У меня есть студент, который хочет понять, что у бактерий является первичным — одна мембрана или две мембраны, — и не может, потому что дерево всё время перестраивается в глубоких узлах, а от этого весь сценарий меняется.
— Не очень понятно, почему.
— Из чисто математических соображений, там просто сигнал пропадает. В зависимости от топологии дерева эволюционные сценарии сильно меняются. А дерево ненадежно, потому что накопилось уже слишком много изменений. В пределе, если прошло бесконечное время, то все окажутся друг другу родственниками на одинаковом уровне.
Но тем не менее масса интересного восстанавливается надежно. Архейная гипотеза — это же началось как анализ последовательностей? Думали, что это такие странные и разные бактерии, а потом Вёзе на молекулярных данных их объявил самостоятельной большой группой. Затем гипотеза симбиогенеза Линн Маргулис о том, что митохондрии — это потомки каких-то бактерий. Она была доказана теми же молекулярными деревьями, и стало ясно, о каких именно бактериях идет речь. И вот теперь, видимо, окончательно доказано, что мы являемся химерой из локиархей, которых только недавно молекулярно вычислили из метагеномов (сначала сделали их геномы, а уже потом культивировали) и альфа-протеобактерий (правда, сейчас Кунин туда хочет еще подмешать вирус как третью компоненту 4 — вот через лет пять и посмотрим, хорошо ли получилось). В общем, это такой чудесный новый мир.
Да, и последнее — это про виды-двойники, которых морфологи не различают.
— Конечно, про это все говорят и признают, что здесь молекулярные методы просто бесценны.
— А потом они находят морфологические признаки.
— Да, потом они находят, если как следует покопаться, всё так. Но вот про то, что какие-то группы «перемещаются» по дереву туда-сюда, тоже говорят. И про то, что с крупными таксонами более-менее понятно, в отдельных группах где-то понятно, а где-то не очень.
— Ну хорошо. Во-первых, это просто означает, что недостаточно данных собрали, и ситуация еще не устаканилась. Это как с китами было — там же тоже первые деревья были не безумно надежны. А потом, когда появилось достаточно данных, оно всё и выстроилось. И когда какие-то группы гуляют туда-сюда, это просто означает, что либо плохо строили деревья, либо действительно данных не хватает, просто надо немножечко подсобрать.

Там может быть другая проблема, с которой только молекулярными методами и справляются. Скажем, если мы про цветочки говорим, то там еще гибридизация бывает. И тогда эволюция получается не очень древесная. Любимый пример, который Мария Логачёва и Алексей Пенин изучают с очередным поколением аспирантов 5, — это пастушья сумка (Capsella bursa-pastoris). Она безумно успешная, растет везде. Но это гибрид двух очень изолированных — во всяком случае, сейчас — видов. Каждый из них гораздо менее универсален, он эндемик, сидит в своем ареале и никуда не вылазит. А пастушья сумка болтается всюду. Но как понять, что она гибрид? Нужно смотреть каждый геном — отцовский, материнский. Дешевых способов рисовать молекулярные деревья с учетом гибридизации я, пожалуй, не знаю. Хотя, может быть, они есть.
А дальше начинаются мои любимые анекдоты. Вот денисовцы — чисто молекулярный объект. Сколько там осталось от этого денисовца? Полтора зуба и три фаланги…
— Не только. Еще челюсть в Тибете нашли.
— Ну да, еще полчелюсти 6. Но с самого начала денисовцы — это чисто молекулярная история 7.

А кроме того, всякие чудесные истории про интрогрессию митохондрий бурого медведя в белого медведя 8, а значит, все белые медведи по материнской линии являются потомками одной бурой мамы-медведицы, которая известно, где и когда жила. И таких историй много на самом деле.
Еще есть истории про инфекционные раки, которые тоже чисто молекулярные 9. Это не вирусы типа папилломы человека или саркомы Рауса у птиц, которые провоцируют рак, а сами раковые клетки, которые оторвались от первоначального организма-хозяина и функционируют как инфекционные агенты. Венерическая саркома собак — это рак какой-то одной собаки, и известно, когда он появился, — дерево нарисовали и по скорости расхождения выяснили, когда жила та несчастная первая собака, которая заболела 10. А лицевой рак тасманийского дьявола — он, наоборот, молодой, и вроде бы их даже два разных, что само по себе удивительно.
Еще чудесная история такого же сорта. У двустворчатых моллюсков в Атлантическом океане есть инфекционный рак. Причем у одного из этих моллюсков рак от моллюска другого вида: раковые клетки одного вида теперь стали инфекционным раком другого вида. Интересно было бы посмотреть на морфологов, которые бы это каким-нибудь способом, кроме анализа последовательностей, выяснили.
Это такой дифирамб молекулярной филогении, которая делается на рисовании деревьев. Но я понимаю, почему классические зоологи испытывают неуверенность, когда при них произносят слово «вероятность»: потому что на естественнонаучных факультетах в университетах очень плохо учат теории вероятности; просто не тому, чему надо. На самом деле это великое достоинство — то, что в этих деревьях встроен механизм оценки их надежности.
Что могут молекулярные деревья?
— Еще такое «но». Морфологи понимают, как в организме всё устроено, что за что отвечает и как это всё работает. А на уровне молекул, спрашивают они, знаем ли мы путь от молекул до функционирования конкретного органа и тем более организма? И когда узнаем?
— Во-первых, интересный вопрос, не иллюзия ли это понимание. И второе, более существенное. Очень долго разные люди говорили, что надо изучать evo-devo, evolution-development. Ну, до сих пор бытует мнение, что онтогенез повторяет филогенез, но мы знаем, что это неправда 11. Геккель, как известно, свои картинки сильно приукрашивал. Но изначальная идея была правильной: различия между взрослыми существами закладываются в эмбриогенезе, а с эволюционной точки зрения это различия чисто молекулярные — на уровне регуляции экспрессии генов. А теперь это можно изучать содержательно, потому что мы можем сравнивать транскриптомы единичных клеток.
— Мы видим, на каком этапе у них начинают работать те или иные гены?
— Да, мы видим, в какой момент какие гены начинают работать. Для одного какого-то гена это и раньше могли, каждый раз в отдельном эксперименте. И, потратив много сил аспирантов, установили генные сети развития дрозофилы, морского ежа, цветка арабидопсиса. А сейчас мы можем делать транскриптомы единичных клеток, поэтому видим пути клеточной дифференцировки.
Вот когда началась интересная история с филогенией? Когда плотность покрытия существующего разнообразия известными геномами достигла некоторого критического уровня. И тогда стали видны всякие нетривиальные вещи.
— Когда примерно?
— В разных группах по-разному, это же очень неравномерно делалось. Но, по-хорошему, качественный перелом — это, видимо, нулевые. Хотя археи, в общем, — тоже качественный перелом, полученный молекулярными методами, а это конец 1970-х. Хорошие же развитийные серии транскриптомов, сначала тотальных, потом отдельных тканей, а теперь одноклеточных — это прямо сейчас происходит. Недавно была работа: сравнили раннее развитие мыши и кролика 12. Прямо на уровне того, как одни типы клеток сменяют другие. Но чтобы делать это совсем хорошо, нужно двадцать грызунов, и вот тогда вы поймете, как развиваются грызуны. И еще двадцать приматов, чтобы сравнивать грызунов с приматами.
— Это, конечно, невероятно круто. Но дилетантский вопрос: а что нам это дает?
— Ну как, это нам объяснит, почему мышь — это мышь, а кролик — это кролик. Хотели же классики понимать, как всё работает? Был уже довольно старый эксперимент 13, который я очень люблю. Взяли ген, который регулирует развитие передней конечности у млекопитающих. По-моему, его функция состоит в том, что он определяет, в какой момент окостеневает хрящ, такой мастер-регулятор этого развития. Взяли этот ген из летучей мыши и пересадили в геном мыши обычной. И ничего не произошло, выросла обычная мышь. А потом взяли регуляторную область перед этим геном, и ее из летучей мыши пересадили в геном обычной мыши. А сам ген оставили как был. И сразу лапка стала длиннее на 15%. То есть сработал не сам ген, а его регуляция.

Млекопитающие по большому счету отличаются друг от друга не набором генов и даже не вариантами генов, а тем, когда они включаются-выключаются. На самом деле на словах это люди понимали довольно давно — на уровне рукомахания. Но полвека это был такой треп, теоретический, а последние лет десять эти вещи можно потрогать руками, смотреть глазами, в компьютер вложить и начать сравнивать. К сожалению, хорошо это делать очень дорого, но становится проще. В первой статье про транскриптомы единичных клеток их было, по-моему, штук триста, и это была статья в Nature. А сейчас в нормальных статьях их тридцать тысяч — или уже триста тысяч.
Можно делать геномы единичных клеток и, например, смотреть филогению нейронов. Какая была бы наивная картина? Ну, вот есть мозг, и в каждом участке мозга нейроны — родственники друг другу, потому что какая-то клеточка делилась, и ее потомки сформировали этот участок. Оказывается, ничего подобного: нейроны, которые находятся в одном месте, генетически друг другу очень далекие родственники. Настолько далекие, что их общий предок мог вообще не быть предшественником нейронов, он был какой-то более ранней клеткой. А их функциональная идентичность определяется тем, в какое место они попали.
Задним числом понятно, что инженерно это очень правильное решение. Почему? Представим себе, что участок мозга — это потомки одной клетки. Это означает, что если в процессе онтогенеза эта клетка случайно умерла, то у вас исчез целый участок, потому что клетки, которые из нее должны были развиться, не появились. А ситуация, когда идентичность нейрона определяется тем, куда он попал, гораздо более устойчива. Потому что даже если какие-то клетки в процессе онтогенеза случайным образом погибают, а это неминуемо, то место их потомков занимают другие. Это гораздо более устойчиво, чем жесткая иерархия.
Есть еще одно полезное приложение клеточной филогенетики — реконструкция истории раковых опухолей. Это те же древесные методы. Можно смотреть, в каком порядке появлялись мутации, как различаются клоны, каково происхождение метастазов.
Таким образом, область применения молекулярных деревьев на самом деле гораздо шире, чем кажется на первый взгляд. Скажем, эмбриология на уровне геномов — т. е. какие клетки от каких происходят. Это можно дополнить транскриптомикой и описывать дифференцировку клеточных типов и тканей. Филогения рака — это такая опрокинутая эмбриология, дедифференцировка, регрессия к ранним клеточным типам.
— А в России есть возможность у кого-то делать транскриптомы единичных клеток?
— В России с единичными клетками работают, хотя мало где. Коллеги из Института биологии гена делали структуру хроматина в единичных клетках дрозофилы; мы с ними потом эти данные обрабатывали — и получилась хорошая статья 14.
Потом — это моя любимая история — получается, что у насекомых с полным превращением в куколке воспроизводится транскрипционная программа эмбриона 15. Не так ярко, как хотелось бы, но немножко получается. Так что evo-devo — то, что люди давно хотели, — теперь можно делать арифметически.
«Вопрос из схоластического превращается в чисто вычислительный»
— Если вернуться к общей биологии. Я всем задавала вопрос: изменились ли критерии вида? Вообще, что считать видами? И было общее мнение, что виды — это, безусловно, реальность, но критерии вида размываются.
— Ну вот неандертальцы — это отдельный вид или нет?
— Ну, есть разные мнения. Но скрещивание происходило.
— Скрещивание-то происходило, но мальчики были дохлые. Оно было успешным, потому что мы его потомки, но при этом оно было не вполне успешным, потому что гибриды мужского пола, видимо, были слабо фертильными, и это мы тоже видим в геномах 16.
— И как из этого ответить на вопрос, отдельный ли это вид?
— Эта проблема была всегда, только она решалась на уровне разговоров. А сейчас для нее возможны некоторые количественные оценки.
— Я и пыталась всех вывести на количественные оценки.
— Есть классической определение, которое я услышал от Алексея Симоновича Кондрашова: вид — это такой набор особей, между которыми происходит свободный обмен генетическим материалом.
— Бывает так, что группы в природе изолированы, и свободного обмена не происходит, но если их «посадить в одно ведро» (не мое выражение), то особи вполне могут размножаться.
— Есть много разных схем экспериментов. Например, вы сажаете вместе самца одного вида и самку другого, и они начинают размножаться, потому что делать им нечего. А если бы у самки был выбор, то она бы взяла самца своего вида, а на того бы внимания не обратила. Это классическая вещь: должен быть эксперимент с соревнованием. Второй вариант. Есть наездники, два разных вида, они совершенно не скрещиваются — не дают потомства. Но они превращаются в один вид, если их накормить тетрациклином, потому что скрещиваться им не дают вольбахии. Вообще, в биологии всегда есть серые зоны. Есть понятные крайности и есть более или менее широкие переходные ситуации. Нормальный биолог к этому относится без невроза.
— Зоологи ищут количественный уровень молекулярных различий, который мог бы служить критерием вида. И в разных группах он разный.
— Совершенно верно. В разных группах он разный, и человек от шимпанзе отличается один раз на 100 букв, а два экземпляра дрозофилы друг от друга тоже отличаются один раз на 100 букв. Значит, с точки зрения дрозофилы, человек и шимпанзе — это один вид. Но потомства не получается, проверено: энтузиасты этим делом сто лет назад занимались 17.
В принципе, можно для каждого семейства собрать людей, которые им занимаются, и они примут решение, что, скажем, у жуков мы выделяем один вид при таком-то сходстве такого-то гена, а у бабочек — при таком-то сходстве другого гена… Но, во-первых, я не очень понимаю, чем это будет полезно, — это всё равно не содержательное, а техническое определение. Во-вторых, всё равно будет куча исключений, потому что в биологии всегда бывает куча исключений, и всё равно таксономисты будут ругаться. В-третьих, это надо будет действительно делать отдельно для каждого таксона, что само по себе, по-моему, очень смешно. Будет толстый справочник: в таком отряде вид — столько-то процентов сходства по такому гену, а в другом отряде вид — другое количество процентов по другому гену. И на каждой странице печать соответствующего департамента.
Правильная подход, но на порядки более дорогой — это секвенировать геномы заметного количества особей того и другого вида, а дальше посмотреть, есть ли поток аллелей туда или сюда. Если потока аллелей не было — значит, это разные виды. Если вы видите, что они много раз гибридизовались и еще продолжают — значит, это один вид, дальше можно делить на подвиды, как хотите, — вот только что такое с дальневосточными тиграми проделали: поделили на подвиды по геномам 18. Понятно, что в подавляющем большинстве случаев для этого не будет денег, но для каких-нибудь важных и находящихся под угрозой исчезновения видов понимать генетическую структуру популяции необходимо, в частности, чтобы правильно планировать защитные мероприятия: сохранять все подвиды, но при этом их не перемешивать.
Возвращаясь к evo-devo. Мы понимаем, что для видообразования существенны изменения в регуляции, которые меняют морфологию и физиологию. Они тоже происходят в результате мутаций, но при этом доля таких мутаций мала. Поэтому определение вида, основанное просто на сходстве последовательностей, даже и концептуально не очень хорошо, потому что меряет время расхождения популяций, но не содержательные различия.
Вот есть классические цихлиды в африканских озерах 19; у них идет взрывное видообразование, и при этом они свободно скрещиваются. То есть свободно они как раз не скрещиваются — не хотят, — но если им не давать выбора, то они будут скрещиваться, и получится предковая форма, серенькая. Поскольку там виды очень молодые, то они генетически, по последовательностям, очень похожи, а морфологически они довольно разные. Опять-таки, это означает, что у них какие-то конкретные гены немножко по-другому работают. Поэтому, например, форма рта получается разная, и в итоге кто-то соскребатель, кто-то — хищник, а кто-то — подбиратель со дна или с поверхности. И окраска разная — это важно, чтобы мальчики и девочки узнавали друг друга, а не других. Но при этом они довольно сильно гибридизуются, это видно.
— Но это не мешает считать их видами.
— А пофиг… Я задавал зоологам другой вопрос. Виды-то ладно, виды мы еще худо-бедно как-нибудь определим. А род — это что такое? И все остальные таксономические уровни? Самые честные зоологи говорят, что да, конечно, остальное — это просто конструкты, которые мы делаем для удобства. С одной стороны, да. А с другой… Скажем, что такое отряд у млекопитающих — это довольно понятно (кроме действительно каких-то особых случаев, как с китами). И понятно, почему понятно. Видимо, потому что они в какой-то момент все очень быстро дивергировали. Отряды млекопитающих схлопываются примерно 70 млн лет назад. Это ситуация, как у соцветия укропа, — одномоментное разделение на много веток, потом еще одно — четкая иерархия. Другой вариант — у рябины: ветвление в соцветии хаотическое, но на выходе всё равно получается зонтик. Поэтому если бы мир был устроен так, как зонтик укропа, то у нас были бы роды, семейства, всё было бы хорошо. Если мир устроен так, как зонтик рябины, то никаких родов и семейств нет, и мы можем только формально сказать, что 50% чего-нибудь — это отряд. Почему 50%? Потому что нам так удобно.
Вот языки, похоже, устроены так, как укроп. Я спрашивал у лингвистов, там действительно есть понятие языковой семьи, оно разумное, семьи примерно одного уровня членения. И нет полноценных гибридных языков (пиджины не в счет).
Вообще говоря, это можно было бы посмотреть, я даже знаю, как. Чем отличается укроп от рябины? Если мы спроецируем узлы ветвления укропа на ось, которая идет вдоль соцветия (то есть на продолжение стебля), то у нас будут точки, в которые проецируется очень много узлов ветвления. А если мы проецируем рябину на ось, которая идет вдоль соцветия, то мы ничего подобного не увидим, потому что ветвления будут хаотически разбросаны по этой оси и никаких сгущений не будет.
Почему отряды млекопитающих хороши? Если мы берем дерево млекопитающих и проецируем его на ось времени, то мы видим очень большое сгущение в тот момент, когда, собственно, сформировались нынешние отряды. И это означает, что отряд млекопитающих — это разумная единица, вполне объективная, с которой можно оперировать.
То есть на самом деле вопрос о том, существует ли таксон высокого уровня содержательно или это чисто технический конструкт без всякого содержания, решается так: берем дерево, проецируем его на ось времени и смотрим, есть ли сгущение.
— С Еленой Темеревой, профессором кафедры зоологии беспозвоночных биофака МГУ, в «Разговорах за жизнь» 20 вы говорили про кембрий: если в кембрии сразу возникло много-много организмов, значит, их можно считать родоначальниками типов.
— Это значит, что типы тоже существуют. Вопрос из схоластического превращается в чисто вычислительный. У нас может не получиться это сделать по каким-то причинам, но хотя бы мысленный эксперимент мы можем поставить.

— У меня еще такой неожиданный вопрос возник. Как вы думаете, зачем растениям такие сложные геномы? Вроде бы им не приходится решать в жизни таких сложных задач, как животным, — движение там, обучение…
— А кто сказал, что эти геномы сложные?
— Ну, первых, они очень большие.
— А у амебы еще больше, что теперь? Большой геном — это нехорошо, большой геном —это означает, что отбор неэффективный. Кроме того, надо смотреть, каких функциональных классов генов у растений много. Скажем, у них более сложный метаболизм, они продуцируют много вторичных метаболитов, потому что их все грызут, а они не могут сбежать, и надо защищаться какой-нибудь химией. И вообще, им нужны гены на все случаи жизни, потому что они съесть никого не могут, значит, надо всё синтезировать, и стрессы надо переносить, потому что не спрячешься.
С другой стороны, сложность в нашем смысле — поведенческие реакции, а для этого не нужно очень много генов. Для этого нужна гибкая система регуляции. Опять же прорастание нейронов — нужно соблюдать какие-то общеинженерные принципы, за счет не очень большого количества генов. А дальше оно само получается.
Теперь формулируется интересный вопрос, ответ на который я не знаю: что будет, если сравнить сложность регуляторных сетей. Может оказаться, что у нас регуляция в значительной степени комбинаторная, и поэтому очень много регуляторных генов не нужно, нужны разные комбинации их продуктов. А у цветочков она может быть плоская, когда на каждое условие нужен свой отдельный регулятор. Это надо просто посмотреть.
1 См. предыдущие интервью: trv-science.ru/tag/molekulyarnaya-phylogeniya
2 Гельфанд М. С. Молекулярная эволюция: как киты уходили под воду // Природа. 2016. № 10. elementy.ru/nauchno-populyarnaya_biblioteka/434242/Molekulyarnaya_evolyutsiya_kak_kity_ukhodili_pod_vodu
3 Михневич Ю. Амборелла волосистоножковая // Элементы, 22.04.2022 elementy.ru/kartinka_dnya/1547/Amborella_volosistonozhkovaya
4 Штерн Б. Новое о происхождении эукариот: кто кого съел. Интервью с Евгением Куниным // ТрВ-Наука № 399 от 12 марта 2024 года.
trv-science.ru/2024/03/koonin-o-proishozhdenii-eukariot/
5 Коровина А. Мария Логачёва: об эволюции растений, создании идеальной гречихи и пангеномной эре в исследованиях // Биомолекула, 24 августа 2022 года.
biomolecula.ru/articles/mariia-logacheva-ob-evoliutsii-rastenii
6 Гельфанд М. Тибетский денисовец //
ТрВ-Наука № 279 от 21 мая 2019 года. trv-science.ru/2019/05/tibetskij-denisovec/
7 Огнёв А. В поисках утраченного генома. Нобелевская премии по физиологии и медицине. Интервью с Михаилом Гельфандом // ТрВ-Наука № 364 от 18 октября 2022 года. trv-science.ru/2022/10/v-poiskax-utrachennogo-genoma/
8 Сернова Н.В., Гельфанд М. С. Истории, прочитанные в митохондриальных геномах: слоны, медведи, люди… // Природа, 2016, № 12. elementy.ru/nauchno-populyarnaya_biblioteka/433986/Istorii_prochitannye_v_mitokhondrialnykh_genomakh_slony_medvedi_lyudi
9 Гельфанд М. С. Инфекционный рак // Природа. 2016. № 2.
elementy.ru/nauchno-populyarnaya_biblioteka/434079/Infektsionnyy_rak
10 Carlo C. Maley, Darryl Shibata. 2019. Cancer cell evolution through the ages // Science 365, 440–441. science.org/doi/10.1126/science.aay2859
11 Гельфанд М. Песочные часы в биологии развития // ТрВ-Наука № 112 от 11 сентября 2012 года. trv-science.ru/2012/09/pesochnye-chasy-v-biologii-razvitiya/
12 Mayshar Y. et al. 2013. Time-aligned hourglass gastrulation models in rabbit and mouse // Cell. Jun 8; 186(12): 2610–2627.e18. doi: 10.1016/j.cell.2023.04.037
13 Cretekos C. J. et al. Regulatory divergence modifies limb length between mammals // Genes & Dev. 2008. 22: 141–151. doi: 10.1101/gad.1620408
14 Ulianov S. V., Zakharova V. V., Galitsyna A. A. et al. 2021. Order and stochasticity in the folding of individual Drosophila genomes // Nat Commun 12, 41. doi.org/10.1038/s41467-020-20292-z
15 Ozerova A.M., Gelfand M.S. 2022. Recapitulation of the embryonic transcriptional program in holometabolous insect pupae // Sci Rep 12, 17570. doi.org/10.1038/s41598-022-22188-y
16 Martin Petr et al. 2020. The evolutionary history of Neanderthal and Denisovan Y chromosomes // Science 369, 1653–1656(2020). DOI: 10.1126/science.abb6460
17 en.wikipedia.org/wiki/Ilya_Ivanov
18 Гельфанд М. Особенности эволюции древних тигров. Интервью с Дмитрием Гимрановым // ТрВ-Наука № 387 от 19 сентября 2023 года. trv-science.ru/2023/09/osobennosti-evolyucii-drevnix-tigrov/
19 Malinsky M. et al. 2018. Whole-genome sequences of Malawi cichlids reveal multiple radiations interconnected by gene flow // Nat Ecol Evol 2, 1940–1955.
doi.org/10.1038/s41559-018-0717-x
20 Гельфанд М. Фантастические гады и как их изучают. «Разговор за жизнь» с зоологом Еленой Темеревой // Naked Science, 13.09.2022.
naked-science.ru/article/interview/fantasticheskie-gady-i-kak-ih-izuchayut

 (4 оценок, среднее: 4,50 из 5)
(4 оценок, среднее: 4,50 из 5)
Реакция двуликого Гугла на главную проблему молекулярной филогении.
Мир устроен как укроп — Результатов: примерно 143 000
Мир устроен как рябина — Результатов: примерно 22 400
The world is structured like dill — Результатов: примерно 8 210 000
The world is structured like a mountain ash — Результатов: примерно 16 900 000
По мнению русскоязычного Гугла мир устроен как укроп, — его англоязычный лик придерживается противоположного мнения.
Похоже, если верен антропный принцип, то укроп предпочтительней.
Подозреваю, что в рамках Вашего подхода, предпочтительней будет конопля.
Я вот тоже заметил, что сушки с маком не надоедают в отличие от остальных ;)
Проверил ваше подозрение.
Мир устроен как конопля — Результатов: примерно 58 900
The world is structured like hemp — Результатов: примерно 119 000 000
Русскоязычный Гугл остался приверженцем укропа, — англоязычный вместе с Вами предпочел коноплю.
«сейчас Кунин туда хочет еще подмешать вирус как третью компоненту [4]»
По-моему, это неверная ссылка. Там речь о вирусах в другом контексте. А о том, что вирус — это третья важная компонента, ничего не говорится. И не «сейчас», а уже давно есть такая идея.
Интересно про простейших. Почему бы не предположить что многоклеточность произошла раньше чем одноклеточность. У бактерий как минимум, но почему бы и не у простейших?
«…многоклеточность произошла раньше чем одноклеточность»
Вы полагаете, — возможны условия, когда многоклеточному организму выгоднее эволюционировать в одноклеточные?
Аналогия из физики: если каплю воды в кастрюле нагреть до кипения, — она эволюционирует в скопление не связанных друг с другом молекул водяного пара.
Но, может быть, Вы имеете ввиду что-то совершенно другое? — поясните, пожалуйста.
И чем может быть полезно такое допущение.